

 English
English文献解读|J Hepatol(33):从乳头状肿瘤到胆道癌的连续致癌性的基因组和转录组特征
✦ +
+
论文ID
原名:Genomic and transcriptomic signatures of sequential carcinogenesis from papillary neoplasm to biliary tract cancer
译名:从乳头状肿瘤到胆道癌的连续致癌性的基因组和转录组特征
期刊:Journal of Hepatology
影响因子:33
发表时间:2025.01.18
DOI号:10.1016/j.jhep.2025.01.007.
背 景
了解癌症中癌前病变的分子和遗传学特征对早期发现、预防和改善患者生存具有重要意义,尤其是对于预后不良的癌症,如胆道癌。既往对肝内外胆管和胆囊乳头状癌前病变的研究统称为胆管导管内乳头状肿瘤(IPN)和胆囊内乳头状肿瘤(ICPN)。IPN/ ICPN的患病率具有地域差异,东亚的发病率(占所有胆管肿瘤的10-38%)高于西方国家(7-12%)。胆管树乳头状肿瘤,包括导IPN和ICPN,是公认的癌前病变。然而,潜在的连续癌变的遗传特征仍然不清楚。
实验设计
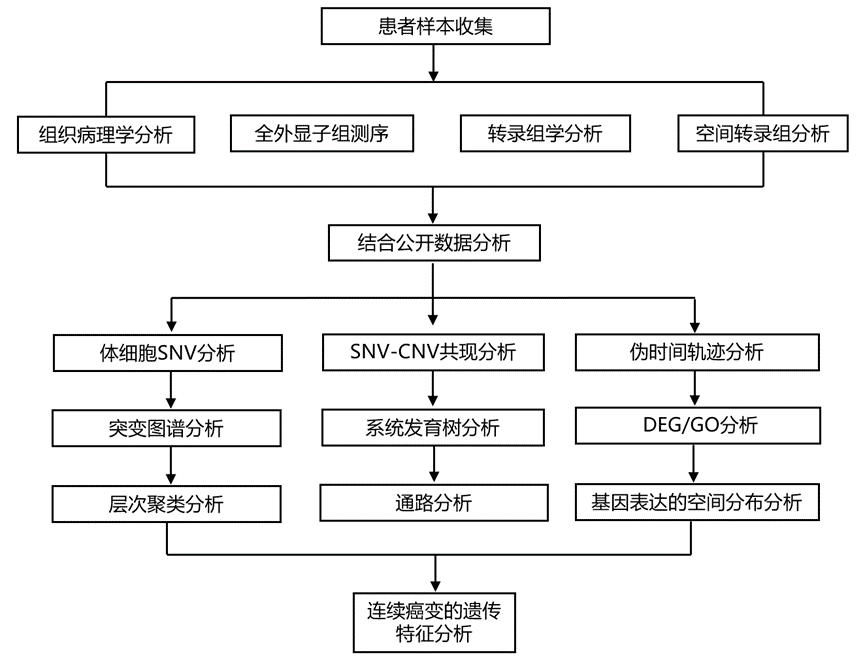
结 果
01

IPN/ICPN的临床病理特征
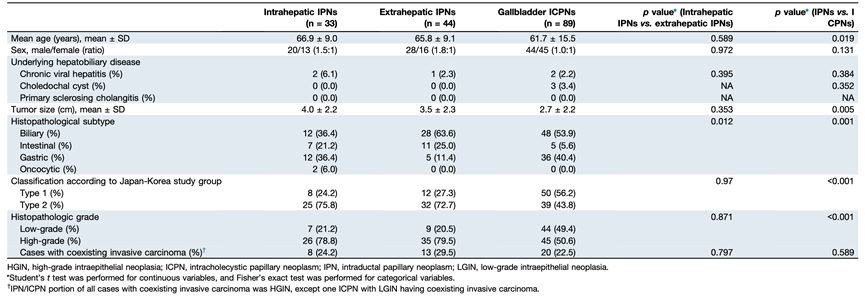
研究团队共筛选出 166 例肝内胆管内乳头状瘤/肝外胆管内乳头状瘤患者,其中包括 33 例肝内乳头状瘤、44 例肝外乳头状瘤以及 89 例胆囊乳头状瘤(图 1A,表 1)。其中,有 41 名患者伴有侵袭性癌变,包括 8 例肝内癌、13 例肝外癌和 20 例胆囊癌。所有侵袭性癌在手术切除时均为局部疾病,无远处转移或扩散迹象。仅有少数患者存在肝胆系统基础疾病,包括 5 例慢性病毒性肝炎患者和 3 例胆总管囊肿患者。在这 166 例原位癌/浸润性癌中,60 例(36.1%)归类为低度上皮内瘤变(LGIN),106 例(63.9%)归类为高度上皮内瘤变(HGIN)(图 1B)。大多数癌伴有 HGIN。进一步的分类显示,88 例(53.0%)为胆管型,23 例(13.9%)为肠道型,53 例(31.9%)为胃型,还有 2 例(1.2%)为多形性细胞亚型(图 1A)。根据日本-韩国研究小组的分类标准,70 例(42.2%)和 96 例(57.8%)的原位癌/浸润性癌病例分别归类为类型 1 和类型 2(图 1A)。胆管型和胃型亚型在肝外原位癌(分别为 63.6%和 11.4%)和肝内原位癌(分别为 36.4%和 36.4%)中的比例存在差异(表 1)。与 IPN 病例相比,ICPN 患者年龄更小,肿瘤尺寸也更小(表 1)。ICPN 中肠道亚型和 2 型的比例低于 IPN 病例。他们对这些样本进行了全外显子组测序(WES)、转录组分析(RNA-seq)和空间转录组分析。
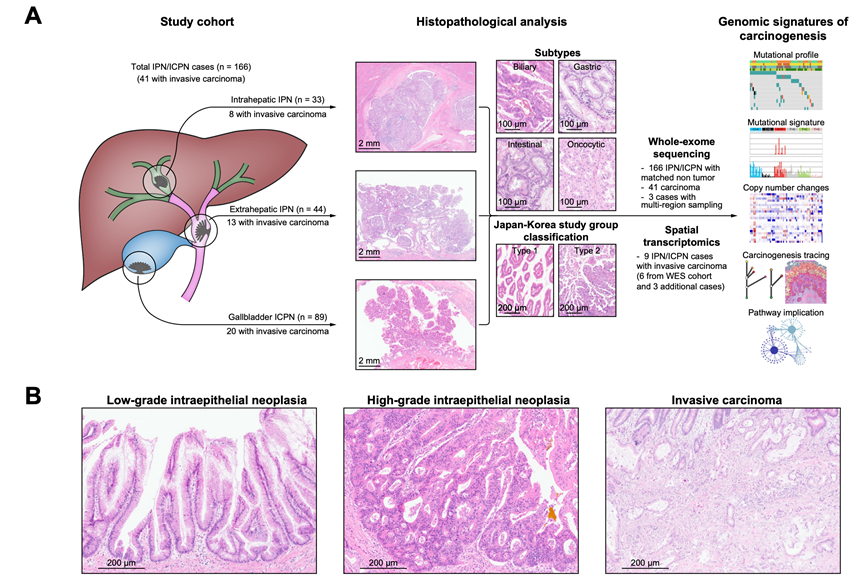
图1. IPN/ICPN的研究概况和组织病理学特征。
(A) 研究队列总结及分析工作。(B) 代表性显微图像。
表1. 研究队列的临床病理特征。
02
IPN /ICPN的基因组图谱
他们从 161 例肝内/肝外胆管内乳头状瘤/胆管内乳头状癌病例(包括 31 例肝内乳头状瘤、43 例肝外乳头状瘤以及 87 例胆管内乳头状癌)中筛选出了 10080 个具有高度可信度的体细胞单核苷酸变异(SNV)。排除了 5 例可能存在读取污染的病例后,没有小的插入-缺失变异通过变异过滤步骤。体细胞变异的发生率在肝内乳头状瘤中高于胆管内乳头状癌。
在致癌驱动基因中出现反复突变的SNV包括 CTNNB1(17.4%)、TP53(16.8%)、KRAS(13.0%)、APC(7.4%)和 SMAD4(6.8%)。TP53(18.2%)、APC(41.2%)和 SMAD4(15.4%)中无义突变的高发生率证实了肿瘤抑制基因的功能破坏。此外,还发现了频繁的拷贝数变异,包括 CDKN2A 缺失(7.4%)、MDM2 扩增(6.2%)和 ERBB2 扩增(6.2%)。在 IPN/ICPN 中主要驱动突变和拷贝数变异的总体频率与之前报道的胆道癌的基因组改变模式相当,但缺少 IDH1 突变这一点除外,因为已知 IDH1 突变在小胆管肝内胆管癌中较为常见。
基因组分析揭示了不同组织位置的肝内胆管癌/肝外胆管癌肿瘤中关键驱动基因的突变频率(图 2)。KRAS 突变主要出现在肝内胆管癌(41.9%)中,而不在肝外胆管癌(16.3%)和肝外胆管癌囊肿(1.1%)中(图 2A-C)。相比之下,SMAD4 主要出现在肝外胆管癌(21.0%)中。在肝外胆管癌中,肝门部胆管癌的 KRAS 突变频率较高,而 SMAD4 突变频率较低,尽管这些差异在统计学上并不显著。CTNNB1 常发生突变,但其突变在肝外胆管癌囊肿中更为频繁(23.0%)高于肝内(12.9%)或肝外(9.3%)胆管癌。在所有组织位置中,HGIN的 TP53 突变频率高于LGIN(在肝内胆管癌中为 0% 对 12.5%,在肝外胆管癌中为 0% 对 37.1%,在肝外胆管癌囊肿中为 0% 对 25.6%)。在类型 2(日本-韩国研究组)中,TP53 突变也比类型 1 更为常见。相比之下,CTNNB1 突变主要出现在低级别胶质瘤伴神经上皮成分(LGIN)中。该突变频率在不同组织病理学亚型(胆管型、肠道型、胃型或嗜酸细胞型)中并无差异。
对已鉴定的SNV进行的通路分析表明,在肝内局灶性结节性增生(IPN)、肝外 IPNs 和肝内钙化性 IPN 中,MAPK 通路(48%)、基因组完整性维持(42%)以及 b-连环蛋白通路(33%)分别是最受显著影响的功能(图 2A-C)。进一步的亚克隆分析确定平均有 2.4 个亚克隆聚类。亚克隆的数量并不因组织病理学亚型、肿瘤分级或解剖位置而有所不同。
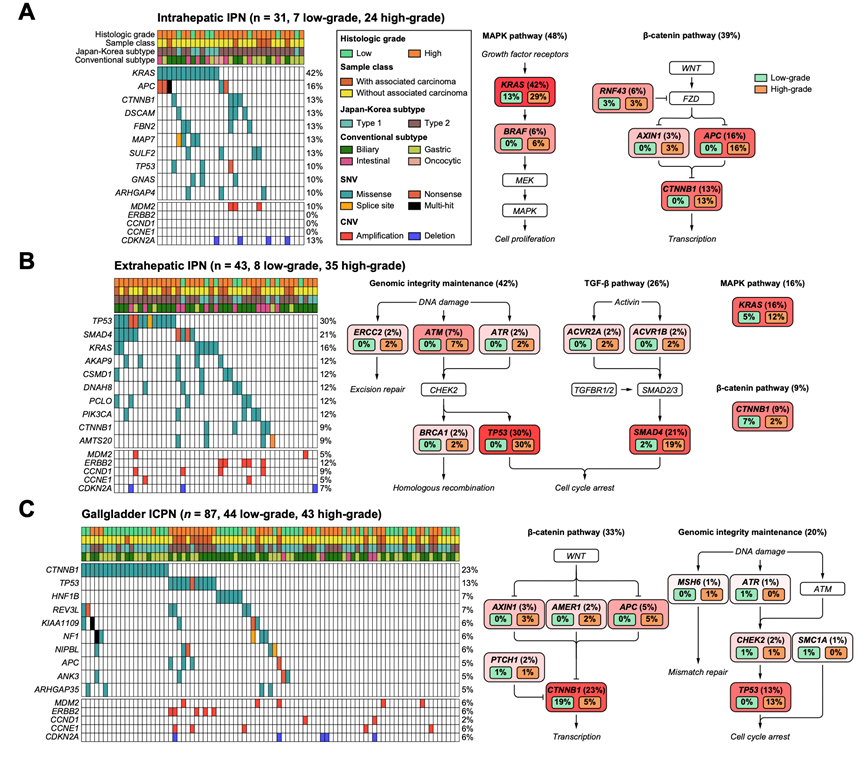
图2. 肝内IPN、肝外IPN和ICPN的突变谱。
(A-C) 左: 前10个突变基因和5个拷贝数在胆道癌中经常改变的基因的癌图。右: 突变通路及各组分基因的突变频率,分别为低级别病变和高级别病变。
03
IPN/ICPN的突变特征
通过全新提取突变特征,发现IPN、肝外 IPN 以及ICPN分别具有两种、三种和四种突变特征(图 3A)。所提取的突变特征归类到先前定义的癌症特征(COSMIC 特征)中,这些特征普遍在所有 IPN/ICPN 中都发现了类似钟形特征(SBS1 和 SBS5);然而,未发现与年龄相关的关联。在肝外 IPN 和 ICPN 中,另外还标注了两个载脂蛋白 B mRNA 编辑酶催化亚基特征(APOBEC)(SBS2 和 SBS13)以及 SBS23(病因不明)。SBS7b(紫外线暴露)仅在 ICPN中观察到。这些特征在侵袭性癌中也有所体现,证实了 IPN/ICPN是癌症前体的证据。
对特征性活动进行无监督的层次聚类分析揭示了样本层面对突变过程的贡献情况(图 3B)。虽然肝内IPN在 SBS1 和 SBS5 的特征性活动中表现出明显的同质性,但在肝外 IPN 和 ICPN 中却观察到了异质性。在 30.2% 和 31.0% 的肝外 IPN 和 ICPN 病例中,观察到了弱至中等水平的 APOBEC 活性,这表明在肿瘤发生过程中存在解剖区域特异性的病因。ICPN 的异质性更为显著,其中还存在 SBS7b 的特征性(29.9%)。基于突变活性的聚类分析证实了肝内 IPN 的病因一致性(图 3C),而与肝外 IPN 和 ICPN 相比具有不同的病因。
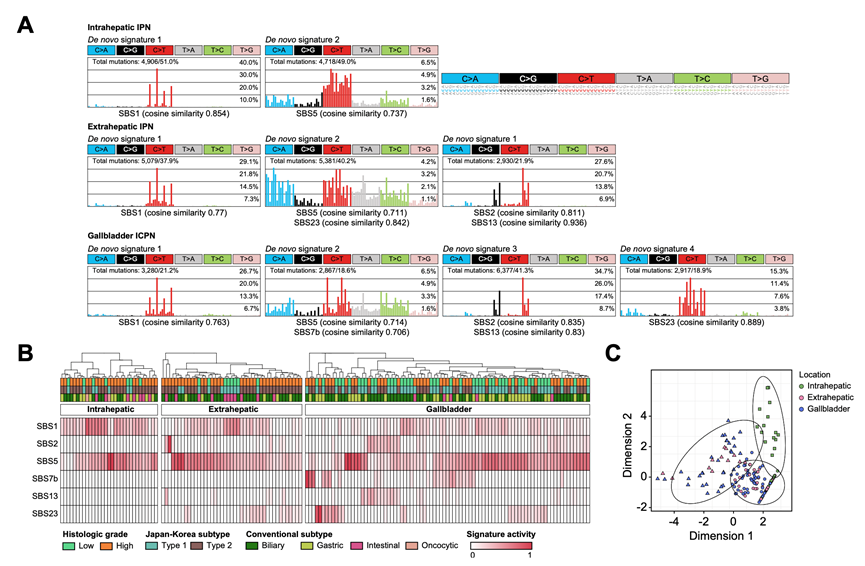
图3. IPN和ICPN的突变特征。
(A) 新发突变特征提取结果。(B) SBS特征热图,为每个解剖位置提供单独的层次聚类。(C) SBS特征提取的K-means聚类结果。
04
IPN/icpn的渐进拷贝数改变
根据异型增生程度(轻度不典型增生、中度不典型增生和癌)对整体的拷贝数变异模式进行分层后,揭示了肝内原发性神经内分泌肿瘤/肝外原发性神经内分泌肿瘤及癌组织中拷贝数改变事件的范围(图 4A)。从轻度不典型增生和中度不典型增生到癌组织,观察到拷贝数变异事件的数量显著增加(图 4B)。此外,在肝内原发性神经内分泌肿瘤/肝外原发性神经内分泌肿瘤中,HGIN 中的拷贝数变异事件数量高于 LGIN。在所有浸润性乳头状瘤/浸润性癌组织以及癌组织中,反复出现的拷贝数变异事件定义为通常涉及的基因区域(图 4A)。染色体增益常见于 1q 和 20q 区域。染色体臂的缺失(5q、9p、17p 和 18q)分别包含了 APC、CDKN2A/B、TP53、SMAD2 和 SMAD4 等基因,这两种组织类型以及癌组织中均检测到了这种缺失。对改变的拷贝数的定量分析进一步表明,从轻度不典型增生到重度不典型增生再到癌组织,其拷贝数逐渐增加(图 4C)。这些结果强调了在轻度不典型增生和重度不典型增生阶段出现的主要拷贝数变异事件的早期发生,并且呈现出向癌组织的逐步进展趋势。
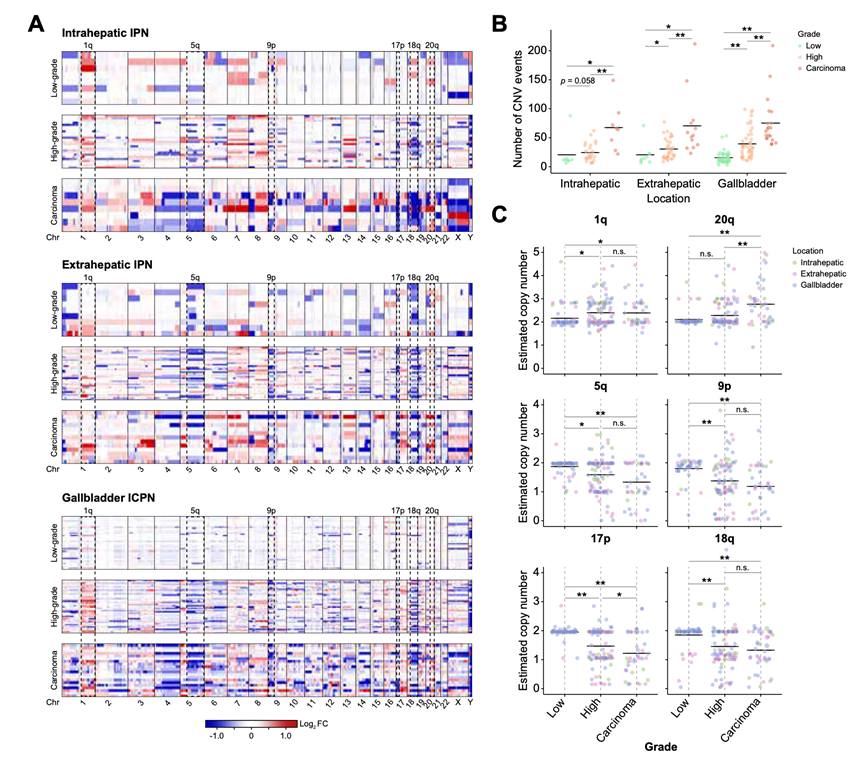
图4. 组织病理分级和解剖位置的全基因组CNV比较。
(A) 拷贝数增益和损失的热图。(B) 散点图显示按组织病理分级的CNV事件数量分布。(C) 染色体臂显著改变的估计拷贝数的散点图。
05
从IPN/ICPN病变发展至相关浸润性癌的连续癌变过程
通过使用贝叶斯概率模型分析SNV和拷贝数变异(CNV)的共现模式,他们阐明了IPN/ ICPN与其相关侵袭性癌变之间的基因组关系(图 5)。在 36 个可分析病例中,32 例(88.9%)的侵袭性癌认定为直接源自 IPN/ICPN。这 32 例源自 IPN/ICPN 的癌变病例(其中 4 例为肝内癌变,13 例为肝外癌变,15 例为胆囊癌变)显示出较高的突变重叠率(平均为 22.0%),其中包括 42 个驱动突变和 4 个拷贝数变异。系统发育分析揭示了从非肿瘤起源到 IPN/ICPN 和癌变在基因组改变方面的进化轨迹(图 5A)。大多数驱动突变为同时属于 IPN/ICPN 和癌变的祖先突变,这表明在 IPN/ICPN 中已经预先建立了主要的致癌环境。然而,这些病例中预癌变阶段的估计持续时间(基于干细胞长度)以及恶性病变开始后的时长(基于分支长度)差异显著,反映了基因组进展的随机性(图 5A)。
三个符合多区域采样条件的病例为进化关系的研究提供了更深入的见解(图 5B)。在病例 30(肝内局灶性肝内胆管上皮不典型增生)中,分析了一个高危病变区域和两个癌灶(癌灶中心:紧邻胆管上皮不典型增生区域;癌灶边缘:距离胆管上皮不典型增生区域更远)(图 5)。这三个病变都具有驱动突变 APC p.K1454*;然而,癌灶中心表现出中等程度的分化,并与胆管上皮不典型增生区域具有更多的体细胞突变(46.2%),包括 KRAS p.G13D 和 CREBBP 亚外显子变异。相比之下,癌灶边缘显示出较差的分化程度和较低的突变重叠(7.2%),这表明其分支较早。值得注意的是,这两个癌灶在基因上不同(突变重叠率为 2.0%),这表明两者都是直接从胆管上皮不典型增生区域衍生而来,而非彼此之间衍生。
在病例 31(肝内局灶性肝内神经内分泌肿瘤)中,对两个神经内分泌肿瘤区域(低级别神经内分泌肿瘤和高级别神经内分泌肿瘤)以及两个癌组织区域(癌组织中心和癌组织边缘)的分析显示,存在一个共同的驱动事件(CDKN2A 缺失);然而,神经内分泌肿瘤与癌组织边缘之间的基因相似性较低,这表明其分支时间较早(图 5B)。由于癌组织与高级别神经内分泌肿瘤(16.5%)之间的共有突变率高于与低级别神经内分泌肿瘤(13.5%)之间的比率,这表明高级别神经内分泌肿瘤是癌组织的起源。除了 CDKN2A 缺失外,KRAS、TSC2 和 ERBB2 的三个驱动突变仅存在于神经内分泌肿瘤中心和癌组织中心之间,这表明癌变是从高级别神经内分泌肿瘤发展到癌组织边缘的,而癌组织中心是较晚发生的事件。
在病例 32(肝外局灶性结节增生)中,对三种类型(癌灶中心型、癌灶边缘型和癌灶中部型)的癌组织以及一个 IPN 区域(高度异型增生型)进行了分析(图 5B)。所有区域都存在两种驱动突变(SMAD4 的 p.R361H 突变和 TP53 的 p.R241T 突变)。然而,癌中心(47.3%)、癌中期(41.8%)和癌边缘(21.9%)与 IPN 的基因组相似度表明存在一种连续分支模式:癌边缘首先发生演变,随后是癌中期和癌中心。这三类癌之间缺乏共同突变这一情况进一步证明了从 IPN/ICPN 发生依赖性癌变而非逐步的癌到癌演变的模式。总体而言,这些结果强调了从 IPN/ICPN 发生的连续癌变通常涉及多发性致癌事件,这些事件导致了癌的内在基因组异质性。
根据进化时间分型的体细胞突变用于识别与癌症发生相关的分子通路。在 32 例由 IPN/ICPN 引发的病例中,IPN/ICPN 和癌组织中分别具有 1834 个和 1349 个突变,影响了 1654 个和 1249 个基因。对癌组织特异性突变的基因本体分析突显了循环系统发育、解剖结构形态发生的调控、参与分化过程的细胞形态发生以及对生长因子的反应作为癌症发生中受影响最显著的途径(图 5C-D)。这些通路是癌症发展中的核心机制,暗示了抗缺氧能力、异常分化和不受控制的生长。相比之下,1834 个 IPN 特定突变涉及在癌症发生中不太显著的显著,如神经元的生成、细胞突起组织和神经元分化。
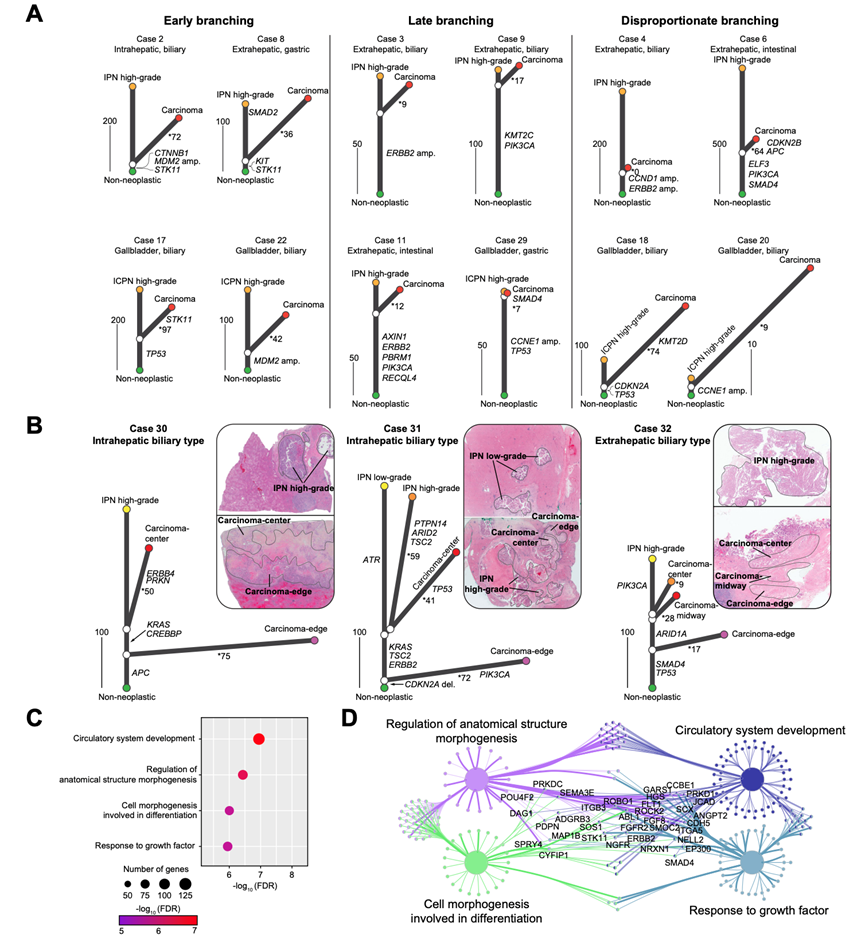
图5. IPN/ICPN进展为浸润性癌的基因组追踪。
(A) IPN/ICPN衍生癌病例的系统发育树。(B) 三个多区域测序病例的系统发育树。(C)GO分析。(D) 通路分析。
06
IPN/ICPN 癌变的转录组特征分析
为了探究IPN/ICPN发生过程中的转录组特征,他们对 9 例伴有侵袭性癌变的 IPN/ICPN 病例(分别来自肝内、肝外和胆囊区域各 3 例)进行了空间转录组分析(图 6)。其中,有 6 例(1 例肝内 IPN:病例 2,2 例肝外 IPN:病例 10 和 32,3 例 ICPN:病例 15、18 和 29)具有匹配的WES数据,并归类为由 IPN/ICPN 引发的癌变。聚类分析确定了具有病理特征的一致性 IPN/ICPN 和侵袭性癌变区域,以及具有最小批次效应的间质特征(图 6A)。
基于均匀流形逼近和投影表示的聚类所推导出的伪时间轨迹分析揭示了癌症发生的关键阶段:起始阶段(称为“根”)、结果阶段(“叶”)以及中间分支点(“分支”)(图 6A)。在 6 例病例(66.7%)中,包括 6 例 IPN/ICPN 源癌症病例中的 5 例以及另外 1 例肝内 IPN 病例(病例 33),轨迹呈现出从 IPN/ICPN 到浸润性癌聚类的线性进展,这与 WES 结果一致(图 6A)。然而,有 2 例(病例 2 和 34)显示了部分相连的轨迹。需要指出的是,轨迹分析的稳健性可能会受到癌症发生过程中径向转录组变化的影响,这使得准确区分 IPN/ICPN癌变特征变得具有挑战性。
对所有九个转录组数据集进行合并后,共识别出六个联合聚类(其中五个对应于浸润性乳头状癌/浸润性癌前癌,一个对应于浸润性癌)(图 6B)。差异表达基因分析(DEG)显示,前 50 个基因在浸润性癌中高度表达。GO分析表明,这些基因在与细胞外基质组织、外源性凋亡途径的调节以及对生长因子刺激的细胞反应的负调控相关的通路中富集,这与基于全外显子测序的通路发现结果大致一致(图 6C)。值得注意的是,一个聚类(标记为橙色的聚类 2)位于浸润性乳头状癌/浸润性癌前癌区域和浸润性癌区域之间,其转录组特征处于中间状态,这表明在癌变过程中存在过渡状态。TERT 表达的空间分布(在恶性转化中起关键作用的因素)在浸润性乳头状癌/浸润性癌前癌和浸润性癌之间没有显著差异。这表明具有高级别上皮内瘤变的浸润性乳头状癌/浸润性癌前癌已经具备了恶性特征,这与本研究的突变数据一致。
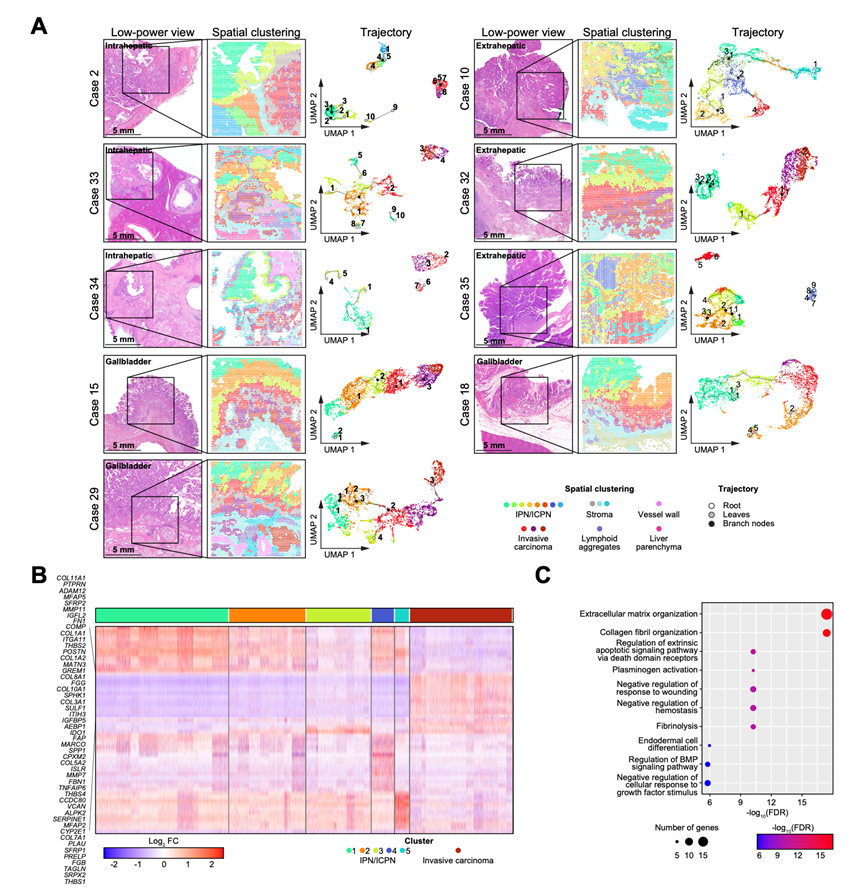
图6. IPN/ICPN与相关浸润性癌的空间转录组学分析。
(A)空间转录组结果。(B) 每个IPN/ICPN或浸润性癌集群从联合聚类中鉴定出的前50个差异表达基因的热图。(C) GO分析。
+ + + + + + + + + + +
结 论
本研究通过多组学分析,发现MAPK(48%)、基因组完整性维持通路(42%)和Wnt/β-catenin(33%)通路突变在肝内IPN、肝外IPN和ICPN中普遍存在。KRAS突变在肝内IPN中富集,而SMAD4突变在肝外IPN中富集(21%,p = 0.005)。ICPN常表现为CTNNB1突变,尤其是在低度病变中。突变特征分析显示,SBS1和SBS5特征在肝内IPN中均匀富集,而SBS1、SBS2、SBS5、SBS13、SBS7b和SBS23在肝外IPN和ICPN中分布不均匀。拷贝数畸变从低级别到高级别上皮内瘤变逐渐增加,最终发展为癌变。系统发育分析显示,89%的癌变源于IPN/ICPN,并通过连续致癌作用而发生,其中大多数IPN/ICPN和癌变之间存在共同的驱动突变。此外,在IPN/ICPN中观察到多灶性、独立的致癌事件,导致突变不同的癌变病变。IPN/ICPN的致癌作用发生在多个亚克隆中,通过突变积累和转录组改变,影响血管发育、细胞形态发生、细胞外基质组织和生长因子反应。
+ + + + +



