

 English
English文献解读|Nat Commun(15.7):肿瘤细胞群落定义了肝癌中肿瘤与微环境的相互依赖关系
✦ +
+
论文ID
原名:Tumor cell villages define the co-dependency of tumor and microenvironment in liver cancer
译名:肿瘤细胞群落定义了肝癌中肿瘤与微环境的相互依赖关系
期刊:Nature Communications
影响因子:15.7
发表时间:2026.02.21
DOI号:10.1038/s41467-026-69797-z
背 景
肿瘤内异质性(ITH)是多种癌症有效干预的主要障碍。单细胞转录组分析(scRNA-seq)已成为研究ITH的热点,研究表明每个肿瘤可能包含多种转录组状态,这些状态由独特的基因表达模式区分。在泛癌单细胞转录组研究中,已定义了多种肿瘤细胞转录组状态,包括细胞周期、上皮间质转化(EMT)以及主要组织相容性复合体(MHC)II相关状态等。尽管这些ITH特征已充分观察到,但不同肿瘤细胞如何在空间上协调运作以及每个肿瘤如何建立其独特的空间格局的机制仍知之甚少。对多种癌症进行点分辨率(每个点包含多种细胞)的空间转录组分析,已开始揭示肿瘤内部的空间格局,在单个肿瘤内观察到基于不同转录组的肿瘤簇的非随机空间分布。在卵巢癌中,基于单细胞空间转录组分析发现,特定的肿瘤细胞状态与肿瘤浸润淋巴细胞在空间上共定位。这些观察结果表明,空间细胞环境在塑造ITH中起着至关重要的作用,并进一步促使他们了解单个肿瘤的空间格局。然而,理解每个肿瘤如何建立其独特的空间格局以及哪些因素驱动肿瘤适应性格局仍然是一项巨大的挑战。
实验设计
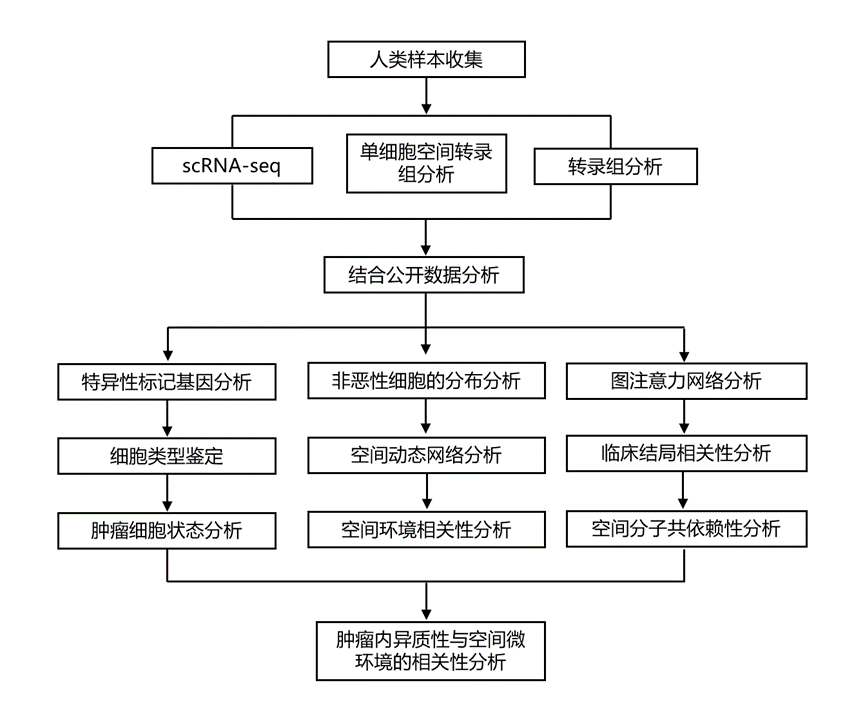
结 果
01

对原发性肝癌患者肿瘤进行单细胞空间表征
为了全面捕捉ITH并研究其在单个肿瘤内的稳定性,研究团队从7例原发性肝癌(PLC)患者的肿瘤内多个部位采集样本(图 1A)。总共从7例PLC患者(4例肝细胞癌,3例肝内胆管癌)的肿瘤核心、肿瘤边界和邻近非肿瘤组织中 采集了50个样本。其中,15个样本(全组织切片)使用CosMx™ SMI原位成像平台进行单细胞空间转录组分析,其余样本在进行了scRNA-seq分析。在本研究中,他们将CosMx™ SMI数据称为“单细胞空间数据”,将scRNA-seq数据称为“单细胞数据”。经过质量控制后,他们获得了2347589个细胞的空间转录组图谱(图 1B)。此外,他们还生成了112506个细胞的scRNA-seq数据(图 1B)。将单个细胞映射到其空间位置时,他们观察到细胞类型注释、细胞类型特异性标记基因的表达以及蛋白质染色之间存在高度一致性,这表明单细胞空间数据能够很好地表征肿瘤图谱(图 1C-E)。
为了进一步确定单细胞空间转录组数据的质量,他们将其与来自同一患者的scRNA-seq数据进行了比较。他们发现两个平台之间的基因表达具有很强的相关性,证实了空间数据的高质量。有趣的是,他们观察到两个平台的细胞类型组成存在显著差异。在scRNA-seq数据中,T细胞占总细胞数的80%以上(图 1F)。相比之下,上皮细胞是单细胞空间数据中的主要细胞类型,这与肿瘤的组织学图像一致。这种差异可能是由于scRNA-seq中的组织解离、保存和文库构建过程所致,在这些过程中,免疫细胞可能优先保留。这些观察结果表明,CosMx平台可能提供肝脏肿瘤的无偏细胞图谱。
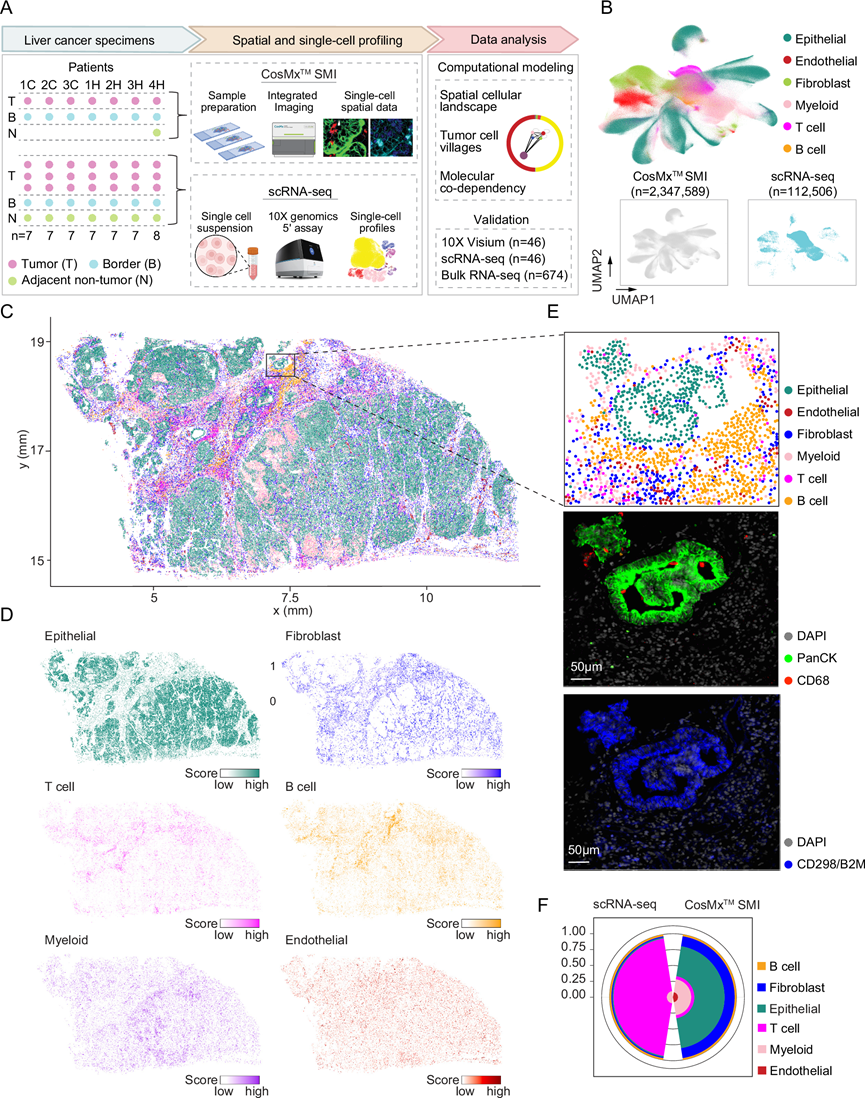
图1. 原发性肝癌的单细胞空间转录组分析。
(A) 本研究的工作流程示意图。(B) 所有分析的单细胞的UMAP嵌入图。(C-D) 代表性肿瘤样本 (1CT) 按细胞类型着色,并显示每种细胞类型的基因评分。(E) 对(C)中选定窗口进行细胞类型注释和蛋白质染色。(F) 比较队列中同一组肝癌患者的基于scRNA-seq和 CosMxTMSMI的细胞类型组成。
02
恶性细胞中转录组的异质性状态
为了解PLC中ITH的程度,他们利用非负矩阵分解(NMF)方法,从单细胞空间数据中确定了所有恶性细胞的转录组状态。使用scRNA-seq数据作为参考,以提高细胞状态识别的可靠性。总体而言,在所有检测的恶性细胞中识别出12种不同的肿瘤细胞状态,包括9种独特的细胞周期、应激反应、免疫反应和运动、金属硫蛋白、EMT、肝细胞样、胆管细胞样、MHC-II、干扰素反应状态,以及3种混合状态:应激/金属硫蛋白、应激/免疫反应和运动以及MHC-II/金属硫蛋白(图 2A-B)。这些状态是基于表达量最高的基因模块及其功能富集情况进行注释的(图 2B)。免疫应答和运动(CCL20、CXCL5、ICAM1)代表了恶性细胞中最常见的细胞状态,突显了肿瘤细胞与免疫细胞之间的相互作用。其他常见状态包括应激/金属硫蛋白、胆管细胞样和细胞周期。有趣的是,他们观察到少量MHC II相关肿瘤细胞。虽然MHC II分子主要由专业的抗原呈递细胞表达,但肿瘤细胞也可以表达这些分子,并且肿瘤细胞的特异性表达与癌症患者的良好预后相关。与恶性细胞相比,非恶性上皮细胞主要富集金属硫蛋白和应激相关程序,这些程序与肝细胞的功能相关。他们发现大多数状态与泛癌研究中鉴定的状态相似,表明不同癌症类型之间存在肿瘤细胞转录组状态的共性。然而,肝细胞样和胆管细胞样状态仅在肝癌中观察到,反映了器官特异性的细胞谱系。值得注意的是,每个肿瘤都包含不同肿瘤细胞状态的混合,突显了每个病灶内显著的ITH。
他们利用单细胞空间转录组数据,通过对每种细胞类型进行基于Harmony的聚类分析,描绘了非恶性细胞的分布图谱。总共鉴定出79个细胞聚类,包括18个内皮细胞聚类、14个成纤维细胞聚类、16个髓系细胞聚类、10个B细胞聚类和21个T细胞聚类,反映了肝癌中免疫细胞和基质细胞复杂且异质的分布图谱(图 2C)。他们进一步基于差异表达基因将这些细胞聚类注释为44种细胞状态(图 2C)。例如,在成纤维细胞群中,鉴定了基质成纤维细胞(COL1A1)、抗原呈递成纤维细胞(CD74、IGKC)、平滑肌细胞(RGS5)、EMT样成纤维细胞(KRT17)、肿瘤样成纤维细胞(VEGFA)、CXCL12 + CAF(CXCL12、IGF2)、IL32+ CAF(IL32、CXCL10)和CCL19+ CAF(CCL19)。类似地,他们也定义了内皮细胞、髓系细胞和淋巴细胞中的不同细胞状态(图 2C)。通过将这些聚类结果与来自个体患者的聚类结果进行比较来验证聚类结果的稳健性,发现大多数来自患者的聚类结果主要映射到特定的基于Harmony的聚类结果,表明Harmony的过度校正程度极低。为了确定从单细胞空间数据和scRNA-seq数据中解析出的非恶性细胞亚型是否一致,他们比较了两种方法得到的每种主要细胞类型的聚类结果。空间数据中的大多数聚类都能与scRNA-seq数据中的特定聚类很好地匹配,这表明尽管空间方法靶向的基因数量少于scRNA-seq,但它仍然有效地捕获了主要的细胞信息。有趣的是,他们在空间数据中观察到一些独特的聚类。例如,在髓系细胞中,TUBB+增殖性肿瘤相关巨噬细胞(TAM)仅在空间数据中发现,这表明单细胞空间方法可能识别出潜在的新细胞聚类。总而言之,CosMx解析的单细胞空间转录组图谱提供了肝癌中非恶性细胞的详细图谱。
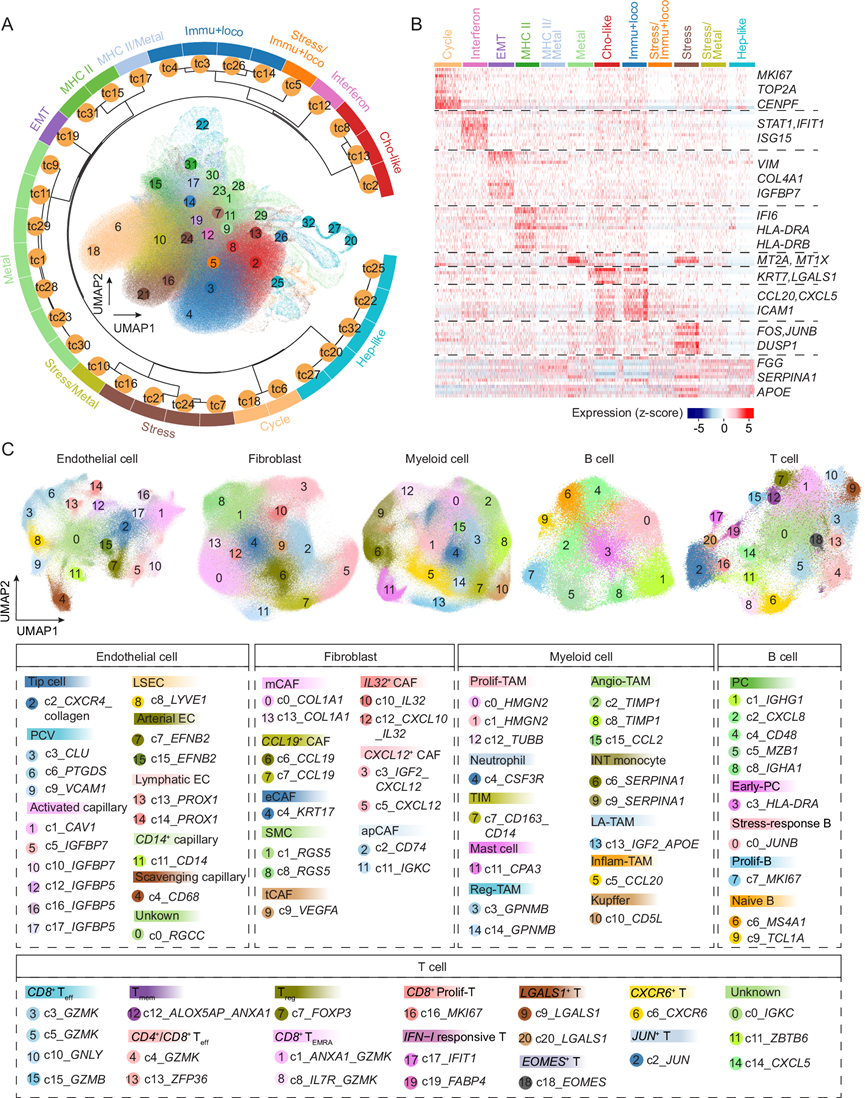
图2. 肝癌细胞的分布图。
(A) 恶性细胞的转录组状态。(B) 与(A)中每种肿瘤细胞转录组状态相关的基因表达谱。(C) 非恶性细胞的UMAP图。
03
通过定义空间动力学网络来表征单个恶性细胞周围的环境
一个成功演化的肿瘤代表着一个空间组织良好的生态系统,其中恶性细胞与其局部环境积极相互作用。这种动态的相互作用不断塑造着肿瘤细胞的功能,从而促进ITH。为了揭示空间环境在驱动ITH中的作用,描绘每个恶性细胞的周围环境至关重要。本研究中解析的单细胞空间数据为此提供了一种独特而强大的方法。他们通过确定每个恶性细胞周围40 µm范围内邻近细胞的组成来表征其周围环境(图 3A)。他们进一步使用Louvain算法,基于周围环境对所有恶性细胞进行了聚类分析(图 3A)。结果表明,属于同一聚类的恶性细胞具有相似的空间邻近细胞。该策略独立于基于转录组的聚类,后者认为特定聚类内的肿瘤细胞具有相似的转录组。通过应用 20 µm、60 µm、80 µm 和 100 µm 的不同半径来测试恶性细胞聚类的稳定性,结果与 40 µm 的结果相似,表明利用邻域信息可以形成稳定的恶性细胞聚类。总共确定了 14 个恶性细胞聚类(对应于 14 种不同的周围环境)(图 3B-C)。他们根据周围环境的组成对其进行了注释,并将每种类型指定为恶性细胞周围的空间动态网络 (SDN)(图 3A-C)。血管化肿瘤是 SDN 的主要类别(SDN-T1 至 SDN-T5),其周围存在内皮细胞。他们还鉴定出一个以肿瘤细胞为主的SDN(SDN-T8),其中恶性细胞构成了周围细胞的大多数。此外,他们还发现了五个富含髓系细胞和成纤维细胞的SDN(SDN-T10–SDN-T14),例如在iCCA样本中观察到的情况(图 3C-D)。这些发现凸显了肝癌中恶性细胞周围微环境的多样性。
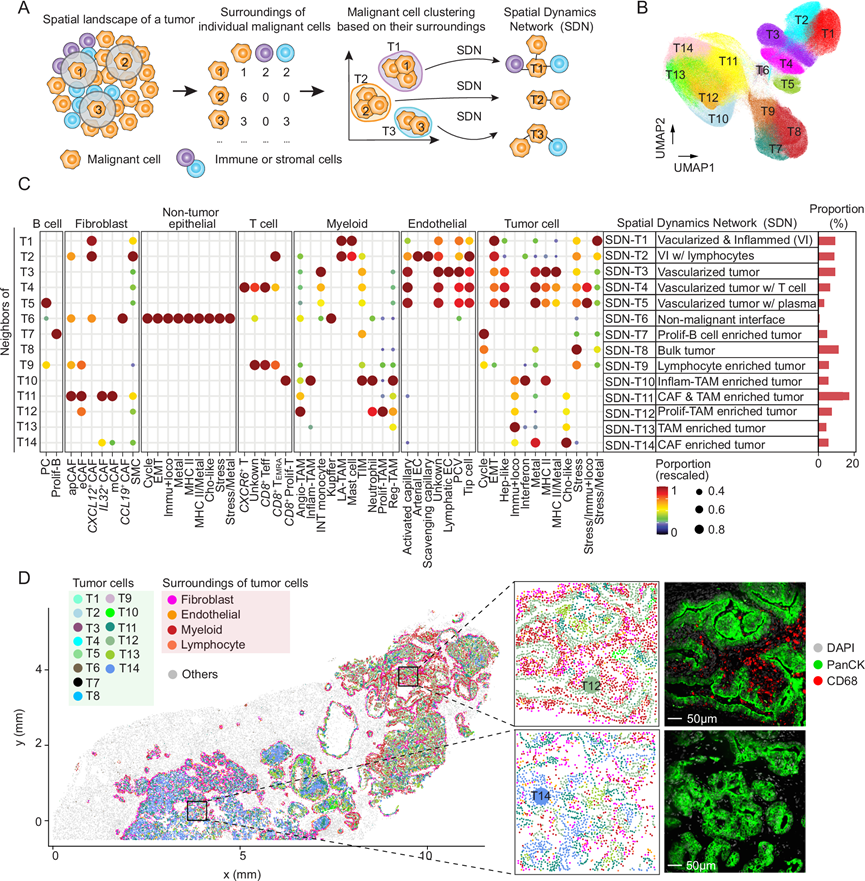
图3. 围绕单个恶性细胞的 SDN。
(A) SDN 确定的示意图。(B)确定的恶性细胞聚类。同一聚类内的恶性细胞具有相似的周围环境特征。(C) 每个恶性细胞聚类周围环境组成的散点图。(D) 展示了一个典型的恶性细胞示例(1CB)(左图),其中恶性细胞根据其SDN进行着色。
04
肿瘤转录组状态与其空间环境相关
为了解空间环境在ITH形成中的作用,他们分析了肿瘤细胞转录组状态与SDN之间的关系。具体而言,采用随机重排策略对恶性细胞状态在SDN上的富集情况进行了分析。值得注意的是,肿瘤细胞状态呈现出非随机的空间分布,每种状态优先富集于特定的SDN(图 4A)。例如,EMT样肿瘤细胞富集于与血管化肿瘤相关的SDN(SDN-T1至SDN-T5)。与此一致的是,已有研究报道EMT转变过程中内皮细胞密度增加。此外,与富集于完全不同SDN的肿瘤细胞状态相比,EMT样肿瘤细胞与内皮细胞的空间距离显著更近(图 4B)。在SDN-T1和SDN-T2中观察到独特的IGF2+脂质相关巨噬细胞(LA-TAM) 群,其中EMT富集最为显著(图3C和4A)。已有研究证实巨噬细胞分泌IGF2以调控EMT。他们还在SDN-T1和SDN-T2中发现了表达IGF2和CXCL12的CXCL1+CAF,提示局部微环境在调控肿瘤细胞EMT表型中起着关键作用(图 3C)。与EMT样肿瘤细胞复杂的局部微环境不同,细胞周期相关的肿瘤细胞在肿瘤细胞占主导地位的SDN(SDN-T7-SDN-T9)中显著富集,而基质细胞和免疫细胞较少。细胞密度分析进一步证实了这一点,细胞周期相关的肿瘤细胞周围的肿瘤细胞密度高于其他肿瘤细胞(图 4C)。此外,他们注意到细胞周期和EMT相关的肿瘤细胞在完全不同的SDN中富集。众所周知,经历EMT的肿瘤细胞增殖能力下降。他们进一步证实了两种肿瘤细胞状态的局部微环境存在显著差异。干扰素是一类细胞因子,能够激活免疫系统清除病原体或肿瘤细胞。在SDN-T10和SDN-T2中,干扰素反应相关的肿瘤细胞富集,而髓系细胞和CD8 +T细胞也存在于这些区域。这些肿瘤细胞表达IFIT1和ISG20的标志物,而这些标志物与抗病毒防御相关。值得注意的是,在本研究队列中,干扰素反应相关的肿瘤细胞主要存在于一名同时感染丙型肝炎病毒和人类免疫缺陷病毒的患者体内,提示该患者可能存在抗病毒反应。在含有成纤维细胞和髓系细胞的SDN(SDN-T11至SDN-T13)中,胆管细胞样肿瘤细胞以及免疫反应和运动相关的肿瘤细胞显著富集。肿瘤细胞状态与其局部微环境之间的配体-受体相互作用分析表明,每种状态下都存在富集的通讯模式,进一步支持了不同肿瘤细胞状态的空间偏好。例如,VEGF介导的相互作用主要见于EMT相关肿瘤细胞与其周围微环境之间。
为了验证肿瘤细胞状态的空间偏好,基于10× Visium平台分析了来自三个公开数据集的肝肿瘤样本。由于10× Visium数据分辨率为点状(直径55 µm),因此针对单细胞空间数据设计的方法无法直接应用于此处细胞状态和SDN的鉴定。为此,通过对单细胞空间数据中相关基因进行平均,构建了每个肿瘤细胞转录组状态和SDN的伪批量基因特征。他们将这些基因特征应用于三个数据集,并采用基于相关性的方法确定了每个肿瘤细胞状态在SDN中的富集情况。他们观察到与单细胞空间数据中发现的富集模式高度一致的富集模式,并且在单细胞空间分析中显示出显著关联的SDN中,肿瘤细胞状态的富集程度远高于其他SDN(图 4D)。总的来说,这些结果表明,肿瘤细胞的转录组状态并非在肿瘤内随机分布。相反,它们与其周围的微环境密切相关,这凸显了空间背景在驱动ITH中的潜在作用。
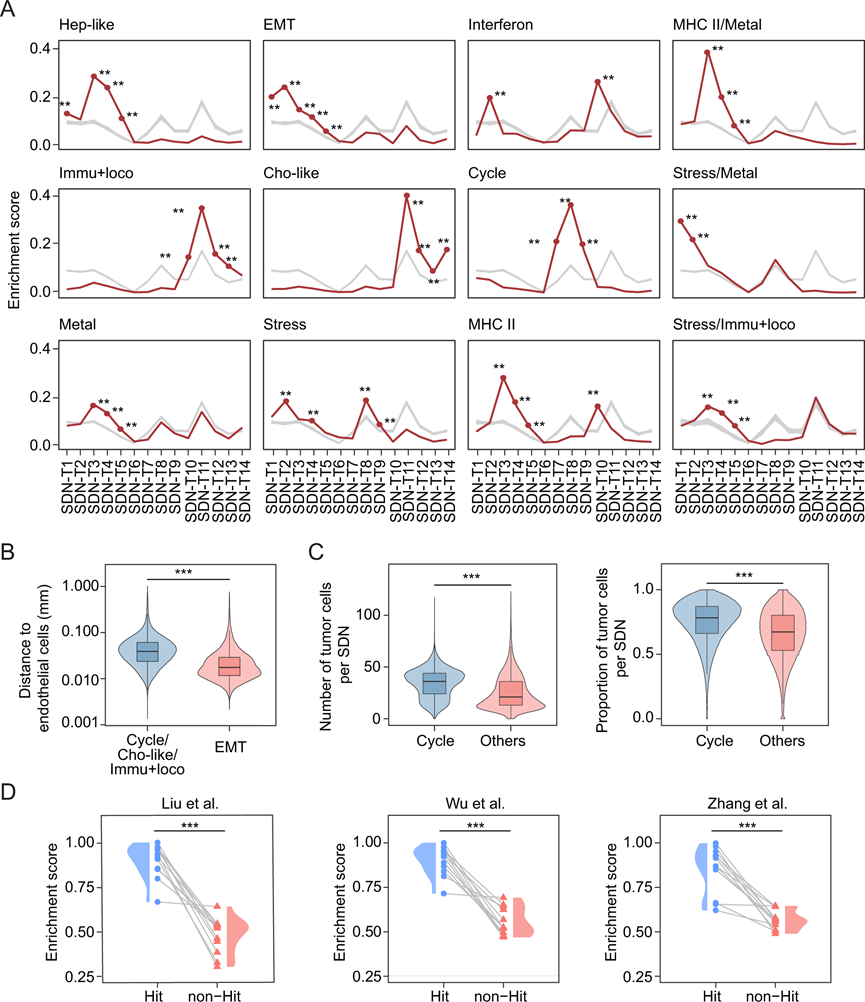
图4. SDN上肿瘤细胞状态的富集。
(A) 各肿瘤细胞状态在SDN上的富集情况。(B) EMT样恶性细胞与内皮细胞之间的距离。(C) 细胞周期相关恶性细胞周围恶性细胞的数量(左图)和比例(右图),并与其它恶性细胞 进行比较 。(D) 结合公开数据验证肿瘤细胞状态的空间偏好。
05
利用图注意力网络(Graph attention networks)鉴定肿瘤细胞群
不同的肿瘤细胞可能在空间上协同作用,共同驱动肿瘤的生长和进展,从而赋予肿瘤生存优势并增强其对干预措施的抵抗力。他们将这种由特定局部环境支持的、不同肿瘤细胞转录组状态的空间协同作用定义为肿瘤细胞“村落”。与人类村庄类似,肿瘤细胞“村落”可能利用共同机制来促进生长和增强防御,从而降低单个细胞的脆弱性。
为了确定肿瘤细胞群落,他们使用了图注意力网络(Graph Attention Networks,GAN),这是一种专门用于分析图结构数据的深度学习神经网络。这些网络利用注意力机制,通过加权相邻节点的重要性来增强特征学习,从而能够捕捉不同肿瘤细胞状态之间复杂的空间关系。通常,将肿瘤内的恶性细胞表示为连通图,其中节点代表由其细胞状态和SDN表征的单个细胞,边连接距离在40 µm以内的节点,这与SDN分析一致(图 5A)。然后,将从图注意力网络导出的嵌入应用于聚类分析,以定义肿瘤细胞村落(图 5B)。排除了三个细胞数量少于5000个的小聚类,它们占恶性细胞总数的比例不到1%。最终,鉴定出八个不同的肿瘤细胞群落(图 5C)。村落1和村落2包含多种肿瘤细胞状态,并具有血管丰富的微环境。相比之下,村落4和村落7主要由细胞周期相关的肿瘤细胞组成,类似于癌症生发中心。村落3、村落5和村落6富含胆管细胞样细胞以及与免疫应答和运动相关的肿瘤细胞,周围环绕着成纤维细胞和巨噬细胞组成的微环境,反映了肿瘤细胞生态系统的完善和资源的充足。村落8主要与免疫应答和运动相关。例如,1CB(一个iCCA样本)中村落3、村落5和村落6位于不同的地理位置(图 5D)。在同一患者的不同区域,肿瘤细胞村落的组成比不同患者之间的组成更为一致。
他们通过差异表达基因分析确定了肿瘤细胞群的替代标记物。所鉴定的基因大多是每个肿瘤细胞群特异性的,并且能够准确预测相应细胞群的存在,表明这些基因可以有效地作为定义肿瘤细胞群的替代标记物。利用这些基因,通过10X Visium平台分析了46例HCC或iCCA肿瘤的空间转录组数据,验证了所鉴定的肿瘤细胞群。为了研究肿瘤细胞群与患者预后的关系,他们对37例HCC或iCCA患者的scRNA-seq数据进行了生存分析,并根据基于替代基因的层次聚类对患者进行了分层。与具有与村落1和2 (V1&2)相关的特征的患者相比,村落 3-8(V3-8)基因特征富集的患者总生存期显著更差(图 5E)。当仅分析HCC患者时,也观察到了类似的趋势。没有单独分析iCCA患者,因为所有iCCA患者主要富集于村落 3-8。使用来自TCGA、LCI(肝癌研究所)和蒙古队列的674例HCC患者的转录组数据,进一步证实了生存关联,观察到肿瘤细胞群与临床结局之间存在一致的关联(图 5F-H)。这些分析突显了肝癌中协调良好的肿瘤细胞群的存在,并表明肿瘤细胞群与患者预后相关。
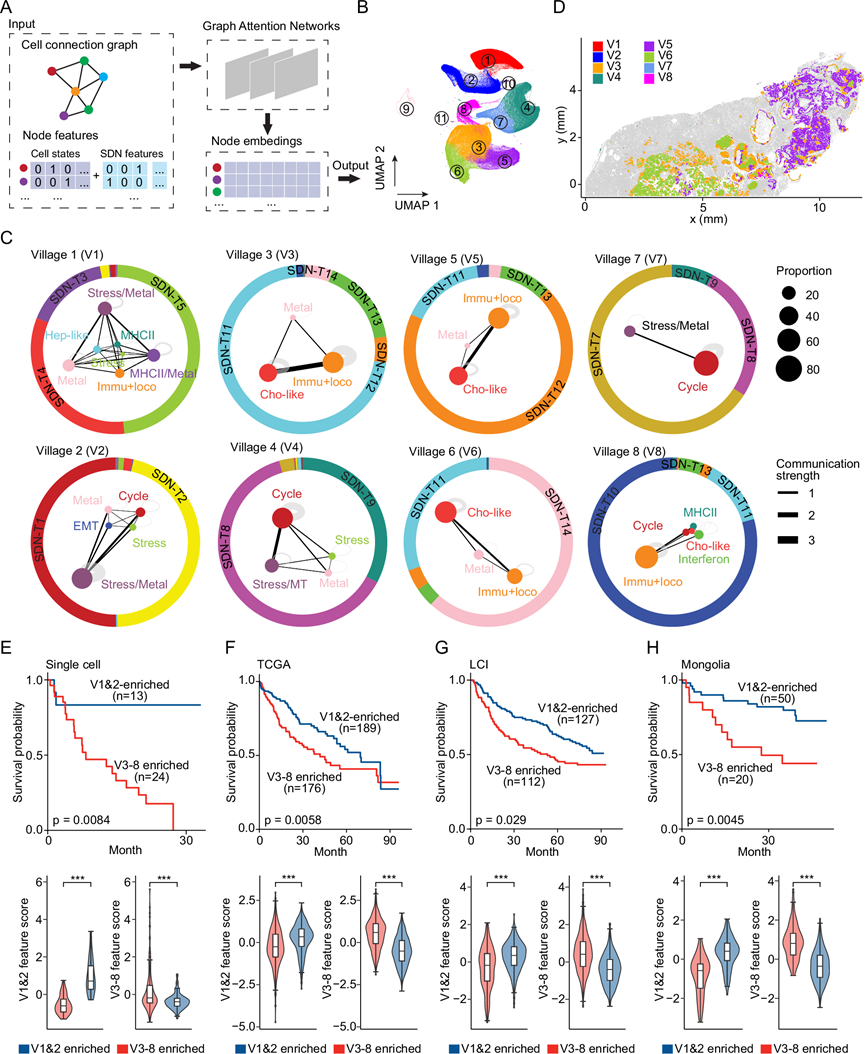
图5. 肝癌中的肿瘤细胞群里。
(A)确定肿瘤细胞群的示意图。(B) 恶性细胞群的UMAP图。(C) 不同肿瘤细胞群的村落图。(D) 肿瘤细胞群的代表性示例(1CB)。(E-H) 上图:在不同队列中,富含V1&2相关特征(蓝色)和V3-8相关特征( 红色)的肝癌患者的总生存期。Kaplan-Meier曲线用于显示各患者组的总生存期。下图:上图中两组患者的村落特征评分。
06
与肿瘤细胞群相关的分子共依赖性
为了揭示驱动每个肿瘤细胞群内细胞协调的关键因素,他们开发了一种评估肿瘤细胞与非肿瘤细胞(基质细胞或免疫细胞)之间分子共依赖性的方法。具体而言,基于40 µm的距离,计算了肿瘤细胞和非肿瘤细胞中每对基因之间的相关性,并假设高度相关的基因对具有很强的共依赖性。利用这种方法,他们鉴定出每个细胞群中相关性最高的基因对,并发现这些基因对具有细胞群特异性,在其各自的细胞群中相关性得分显著高于其他细胞群(图 6A)。通过将空间距离阈值从40 µm增加到80 µm、160 µm、320 µm,以及扩展到细胞群边界之外,进一步评估了每个细胞群中的基因对相关性。值得注意的是,随着细胞空间距离的增加,相关性得分逐渐下降,这突显了基因对关系的空间特异性(图 6B)。为了证明这些基因对关系对每个肿瘤细胞群落的重要性,采用随机森林模型,基于已识别的基因对预测肿瘤细胞所属的细胞群落。当每个细胞群落内肿瘤细胞周围的非肿瘤细胞进行随机打乱时,预测准确率显著下降,与原始细胞群落结构相比(图 6C)。这些发现表明,分子间的相互依赖性可能对维持每个细胞群落的完整性至关重要,而对其的扰动可能会导致细胞群落结构的不稳定甚至崩溃。
在肿瘤细胞中排名靠前的分子中,重点关注了丝氨酸蛋白酶抑制因子Kazal型1(SPINK1),因为它在肿瘤细胞中具有独特的表达(图 6D)。SPINK1相关基因对主要存在于村落6,村落3也有少量检测到(图 6E)。大多数SPINK1相关基因是基质癌相关成纤维细胞(mCAF)相关基因,包括COL1A1、COL1A2、COL5A1、COL4A1、BGN和MGP(图 6F)。使用Visium转录组数据验证了这种空间关系,结果表明SPINK1阳性斑点与成对基因富集斑点之间的距离显著小于随机预期(图 6G)。 SPINK1 和 COL1A1 的多重免疫荧光染色进一步证实了这两种蛋白在 3C 患者中的共定位,该患者在所有患者中村落 6 的比例最高。基因集富集分析表明,SPINK1+肿瘤细胞与转移相关特征相关,而SPINK1-肿瘤细胞则富集于代谢和凝血相关通路(图 6H)。此外,单细胞转录组分析表明,SPINK1+肿瘤细胞通过配体-受体相互作用与 mCAFs 的结合比其他 CAF更为广泛(图 6I)。SPINK1本身鉴定为这些相互作用中的配体之一,其受体为 EGFR。与其它 CAF 相比, mCAF 中SPINK1下游基因的表达水平显著更高。同样,mCAF中 EGFR下游基因的表达也显著上调。当通过随机打乱非肿瘤细胞来扰乱SPINK1及其配对基因之间的关系时,他们发现随机森林模型对村落的预测准确率与原始村落配置相比显著下降,且村落 6 的下降幅度大于村落3,这与 SPINK1 及其配对基因的富集程度较高相一致。在村落6中发现了SPINK1相关的基因对(图 6J)。当这些村落中所有已识别的相关基因均受到干扰时,预测准确率进一步下降(图 6J)。这些观察结果表明, SPINK1阳性肿瘤细胞与mCAF之间的分子共依赖性可能在维持村落3和村落6的完整性方面发挥重要作用。总而言之,这些发现表明,已定义的肿瘤细胞村落可能揭示了每个村落内细胞间独特的分子共依赖性,这为开发旨在破坏这些共依赖性以破坏肿瘤稳定性的治疗策略开辟了道路。
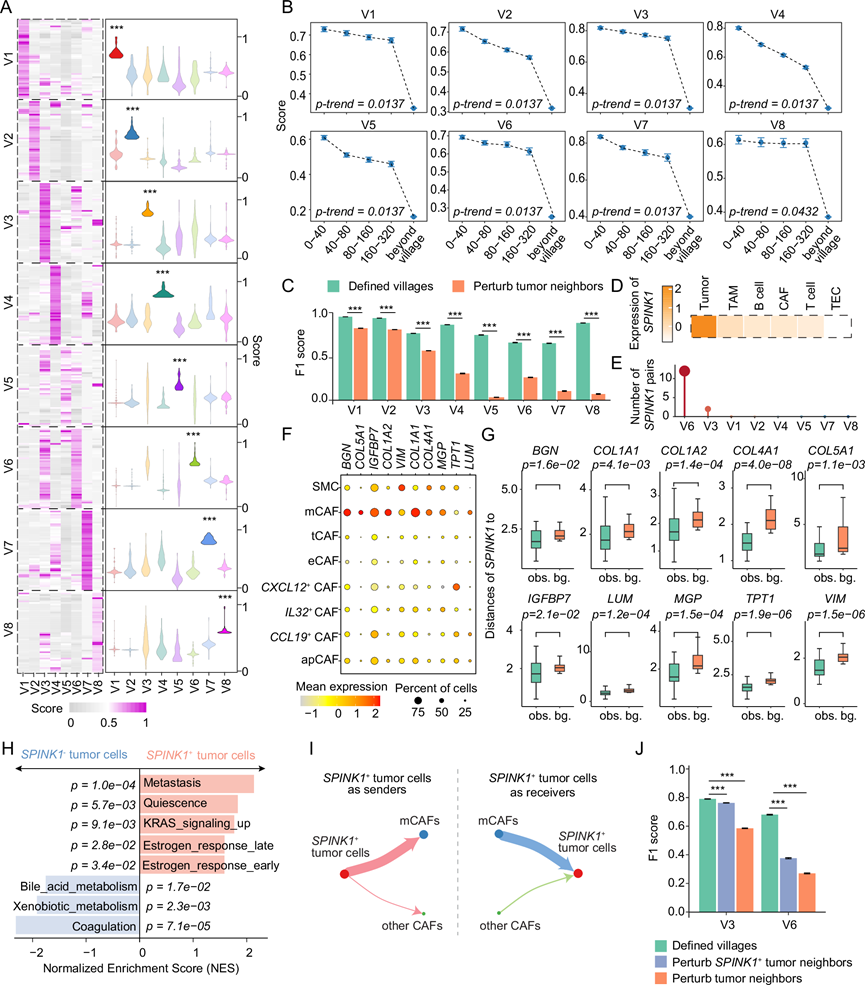
图6. 单个肿瘤细胞群中的空间分子共依赖性。
(A) 每个肿瘤细胞村中前50个基因对的相关性得分。(B) 肿瘤细胞与其邻近细胞之间相关性得分随空间距离增加而变化的梯度。(C) 随机森林模型在已定义的肿瘤细胞村以及扰动肿瘤细胞邻居后的性能的 F1 分数。(D) SPINK1在不同细胞类型中的表达。(E) 每个肿瘤细胞群中SPINK1相关基因对的数量。(F) 不同成纤维细胞亚型中基因的表达。(G) SPINK1阳性点与特定标记阳性点( BGN、COL1A1、COL1A2、COL4A1、COL5A1、IGFBP7、LUM、MGP、TPT1和VIM )之间的距离。(H) SPINK1+肿瘤细胞与SPINK1-肿瘤细胞的基因集富集分析。(I) SPINK1+肿瘤细胞(红色)与 mCAF(蓝色)或其他 CAF(绿色)之间的细胞间相互作用网络。(J) 随机森林模型在已定义的肿瘤细胞群、扰动SPINK1+肿瘤细胞的邻居以及扰动所有肿瘤细胞的邻居时的性能的 F1 分数。
+ + + + + + + + + + +
结 论
本研究利用空间单细胞成像和scRNA-seq,分析了来自50个肿瘤生物样本的超过200万个细胞,开发了一种基于深度学习的策略,用于空间映射肿瘤细胞状态及其周围环境结构,并发现不同的肿瘤细胞状态可以组织成不同的聚类,或称“村落”,每个村落都由独特的微环境支撑。值得注意的是,肿瘤细胞村落表现出肿瘤细胞与其微环境之间村落特异性的分子共依赖性,并且与患者预后相关。通过随机空间重组微环境来扰乱分子共依赖性会导致相应村落的不稳定。本研究使用来自740名肝癌患者的单细胞、空间和整体转录组数据验证了这些发现。这项研究为理解肿瘤空间格局及其对肿瘤侵袭性的影响提供了新的见解。
+ + + + +



