

 English
English文献解读|Nat Cancer(28.5):鼻咽癌的多组学特征分析揭示了不同亚型对诱导化疗的反应特征
✦ +
+
论文ID
原名:Multi-omic characterization of nasopharyngeal carcinoma delineates the subtype-specific landscape of response to induction chemotherapy
译名:鼻咽癌的多组学特征分析揭示了不同亚型对诱导化疗的反应特征
期刊:Nature Cancer
影响因子:28.5
发表时间:2026.04.15
DOI号:10.1038/s43018-026-01149-8
背 景
鼻咽癌(NPC)是一种高度侵袭性的恶性肿瘤,发生于头颈部组织。它在中国南方、东南亚和北非地区尤为常见。目前开展的随机III期临床试验表明,吉西他滨联合顺铂诱导化疗(GP-IC)序贯同步放化疗(CCRT)可显著提高局部晚期鼻咽癌(LA-NPC)患者的生存率,因此该方案已成为LA-NPC的标准治疗方案。然而,仍有20%~30%的患者会出现疾病复发。新出现的证据表明,在放化疗中加入程序性死亡蛋白1(PD-1)或其配体(PD-L1)的阻断治疗可改善LA-NPC患者的临床疗效。然而,目前尚不清楚哪些患者亚组能从中获益最大。目前,临床分期是鼻咽癌治疗选择的主要指导原则。已有的生物标志物,例如EB病毒(EBV)DNA水平和PD-L1表达,对鼻咽癌免疫化疗疗效的预测价值仍然有限。因此,迫切需要寻找能够区分鼻咽癌患者对GP-IC及其他免疫疗法反应差异的新型生物标志物。
实验设计
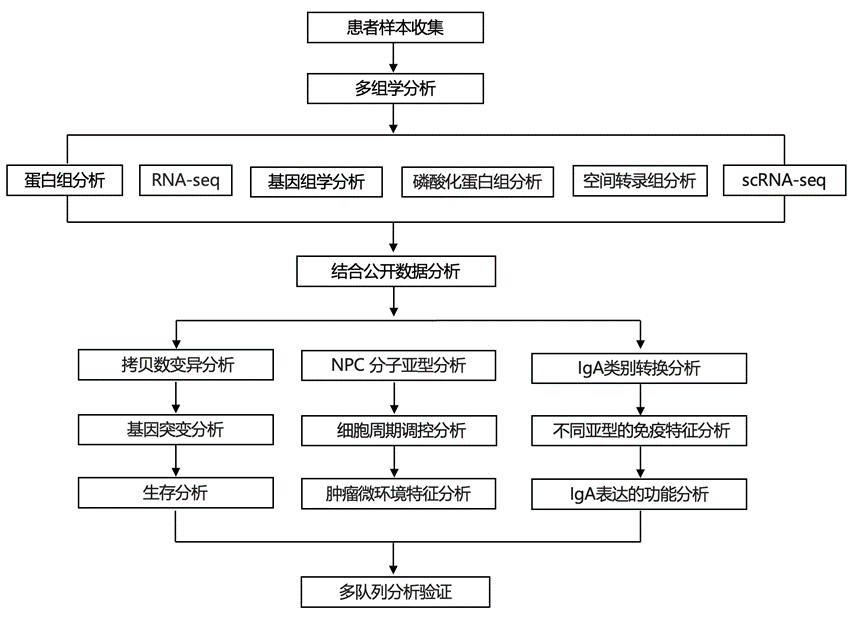
结 果
01

多组学改变与鼻咽癌中GP-IC反应相关
为了探索与NPC中GP-IC疗效相关的生物学和分子特征,研究团队利用来自“中山大学肿瘤协作组-GP队列”(队列1)(图1a)的240例初治NPC患者的样本,评估了其多组学特征。这些患者分别接受了GP+CCRT(n =120)或CCRT(n =120)治疗。他们从每例活检样本中同时提取总DNA、RNA和蛋白质,分别用于全外显子组测序(WES)、转录组分析(RNA-seq)、蛋白质组学和磷酸化蛋白质组学分析。最终,他们获得了225例肿瘤的WES和RNA-seq数据(图1a)。他们通过WES鉴定出14794个突变和25988个拷贝数变异(CNA),并从RNA-seq数据中鉴定出19426个基因。采用数据非依赖性采集(DIA)模式进行的全局蛋白质组学分析定量了来自231个肿瘤的12278个独特蛋白质,而采用数据依赖性采集(DDA)模式进行的磷酸化蛋白质组学分析则从240个肿瘤中鉴定出37166个磷酸化位点(图1a)。
接下来,他们使用包含交互项的Cox回归模型,鉴定了与GP+CCRT方案疗效相比单独使用CCRT方案疗效相关的多组学改变。与之前的研究结果一致,他们发现了一些复发性突变基因和显著基因突变(SMG),这些突变发生在已知的DNA损伤反应基因肿瘤蛋白P53(TP53)和核因子κB(NF-κB)通路基因中,例如TNF受体相关因子3(TRAF3)。在复发性拷贝数变异(CNA)事件中,缺失最常见于3p21.31、14q32.33和11q23.3区域,而扩增则常见于12q14.3、18p11.1和11q13.3区域。在筛选所有复发性基因突变或SMG后,他们仅发现三个突变与GP+CCRT治疗后无病生存期(DFS)和总生存期(OS)获益降低相关(图1b)。值得注意的是,没有SMG与GP-IC疗效相关。11q23.3和1p36.11缺失与GP-IC疗效改善相关,而9q24.1扩增与GP+CCRT治疗组中较高的治疗失败风险相关(图1c)。
接下来,他们筛选了与GP-IC反应相关的单个信使RNA(mRNA)和蛋白质。总共有18个mRNA和54个蛋白质与GP+CCRT治疗组的DFS结果密切相关,而27个mRNA和52个蛋白质与OS结果相关(图1d-e)。与GP化疗反应相关的mRNA改变与失巢凋亡和抗菌体液反应存在一定程度的相关性。然而,每个通路中鉴定的mRNA不足5个,限制了其进一步研究的潜力。同时,与GP-IC反应相关的蛋白质改变显著富集于适应性免疫反应通路,包括适应性免疫和免疫球蛋白生成,突显了适应性免疫在GP治疗反应中的作用。
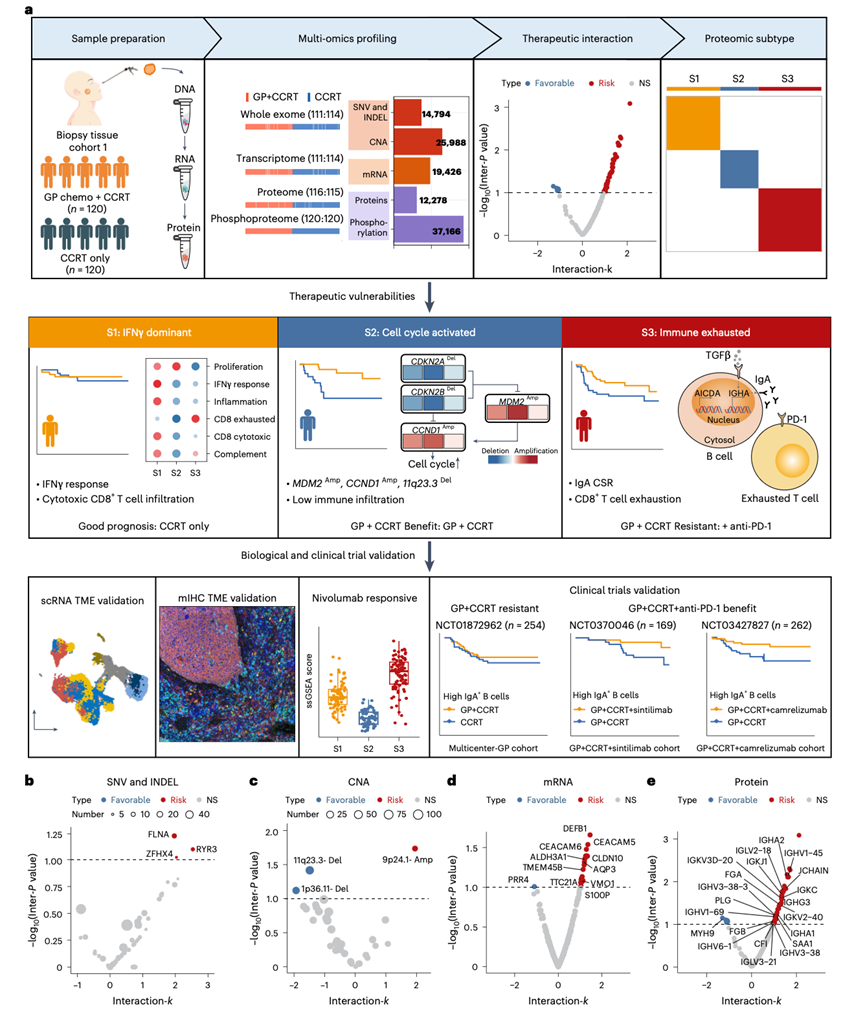
图1. GP-IC 研究设计概述及与生存获益相关的多组学改变。
(a) 多组学研究设计概述。(b-e) 火山图分别描绘了 治疗组与突变、拷贝数变异(CNA)、mRNA和蛋白质交互作用项的k 值和DFS交互作用P值。
02
NPC亚型具有独特的特征,并可从GP-IC中获益
由于蛋白质组学分析揭示了与GP反应相关的大部分特征,他们进一步对蛋白质组学数据进行了无监督共识聚类,随后进行多组学整合,以表征聚类异质性和驱动事件。将肿瘤有效分为三个亚型:S1(n = 80,34.6%)、S2(n = 57,24.7%)和S3(n = 94,40.7%)(图2a),它们表现出不同的免疫学、基因组学和临床特征。S1肿瘤在蛋白质组学、转录组学和磷酸化蛋白质组学层面均表现出与先天性和有效免疫特征(I型或II型干扰素(IFN)、抗原呈递和T细胞活化)相关的最强免疫特征表达(图2a-b)。S2肿瘤表现出细胞周期、增殖和有丝分裂通路显著富集(图2a-b),同时免疫评分最低,但基因组不稳定性最高:肿瘤突变负荷和基因组改变比例升高(图2c-e)。具体而言,S2 亚型在细胞周期相关基因[例如细胞周期蛋白 D1 (CCND1) 和 MDM2 原癌基因 (MDM2)]中拷贝数增加最为频繁,而细胞周期蛋白依赖性激酶抑制因子 2A (CDKN2A) 的拷贝数丢失最为严重(图2a)。S3 肿瘤具有适应性免疫特征,包括 B 细胞增殖和分化、T 细胞受体以及跨所有组学层面的共刺激信号传导(图2a-b)。
临床上,三个亚组在临床分期、无病生存期(DFS)(图2f)和总生存期(OS)方面均表现出显著差异。S1 期肿瘤患者预后最佳,仅接受同步放化疗(CCRT)即可获得充分获益(图2f-h)。虽然S2期肿瘤患者具有侵袭性表型,但GP联合CCRT治疗仍显著改善了预后中等的S2期肿瘤患者的无病生存期(DFS),并略微改善了总生存期(OS)。预后最差的S3期肿瘤患者从GP-IC治疗中获益有限( 图2j)。因此,蛋白质组亚型表现出不同的临床结果和对 GP-IC 的不同反应。
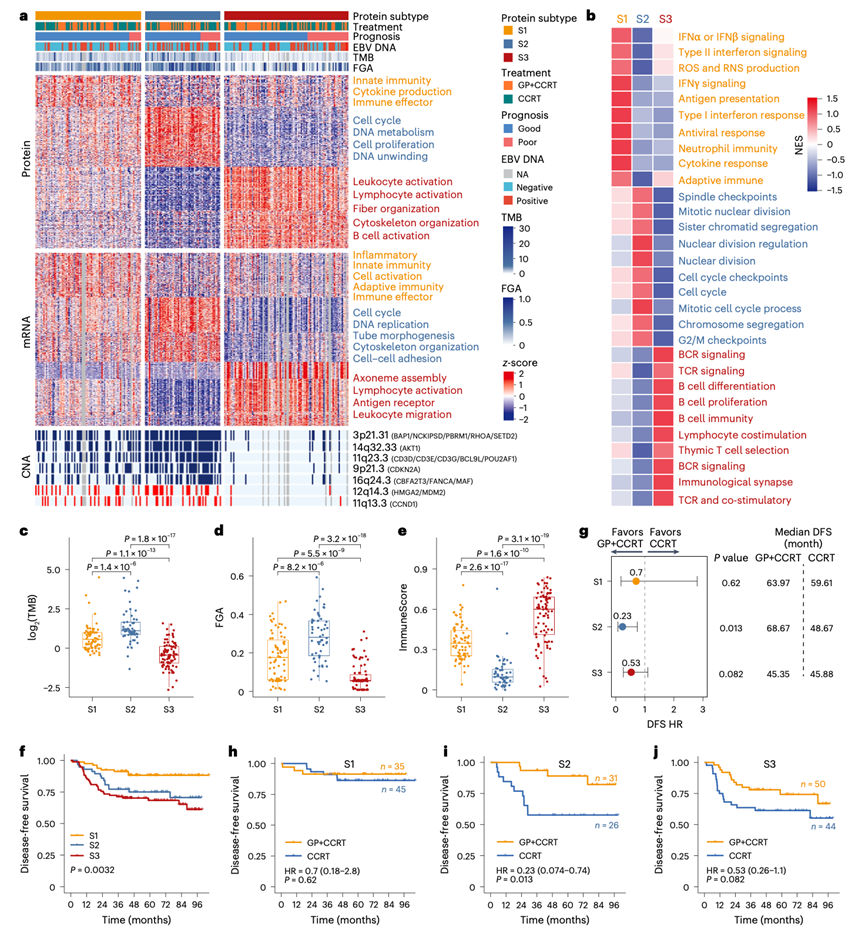
图2. 基于蛋白质组学改变的 NPC 分子亚型及其从 GP + CCRT 中获得的独特益处。
(a) 热图显示了鼻咽癌(NPC)各蛋白质组亚型中特征蛋白、mRNA和拷贝数变异(CNA)的改变。(b) 热图显示了各亚型中显著富集的基因集的标准化富集评分(NES)。(c-e) 箱线图分别描绘了各亚型的肿瘤突变负荷(TMB)、基因组改变分数(FGA)和免疫浸润评分。(f) 不同亚型NPC患者的无病生存期(DFS)Kaplan-Meier曲线。(g) 森林图描绘了各亚型中GP+CCCRT与CCRT的DFS风险比。(h-j) Kaplan-Meier曲线。
03
细胞周期顺式激活驱动GP-IC在S2期发挥益处
为了鉴定GP-IC反应的基因组调控因子,他们分析了CNA对mRNA和蛋白质的影响。大多数重叠的顺式作用事件富集于DNA生物合成以及RNA和蛋白质定位通路。S2肿瘤中CNA-RNA-蛋白质级联基因在细胞周期、DNA代谢和复制通路中富集程度最高(图3a),并频繁出现9p21.3 / CDKN2A / CDKN2B缺失(64.3%)、11q13.3 / CCND1扩增(30.4%)和12q14.3 / MDM2扩增(41.4%)(图3b-c)。总体而言,78.6%的S2肿瘤携带细胞周期相关的CNA,而其他肿瘤中这一比例为31.9%(图3c)。S2肿瘤中CCND1和MDM2的CNA与mRNA和蛋白丰度显著相关,驱动细胞周期激活(图3d-e)。RB1是G1/S期细胞周期的负调控因子,其磷酸化在S2肿瘤中更为丰富,表明其功能失活并导致E2F转录因子释放(图3f)。同时,编码细胞周期蛋白依赖性激酶和E2F5的其他细胞周期基因在S2亚型中也上调。临床上,RB1磷酸化水平升高的患者从GP+CCRT治疗中获益显著。鉴于CCND1 / MDM2扩增驱动 CDK4/6 介导的细胞周期失调,他们发现 CDK4/6 抑制剂(阿贝西利)在模拟 S2 肿瘤的CCND1 / MDM2扩增模型中与顺铂/吉西他滨具有协同作用(扩展数据图4f)。此外,S2 亚型中细胞周期通路激活(图3g)。细胞周期评分高的患者从 GP-IC 治疗中获益显著(图3h)。综上所述,拷贝数变异的顺式效应驱动细胞周期激活,并赋予 S2 亚型化疗敏感性。
在1278个癌症基因普查(Cancer Gene Census)基因中,有25个基因表现出显著的CNA-mRNA顺式调控,其中包括细胞周期调控因子,例如MDM2、CCND1和CDKN2A。值得注意的是,11q23.3缺失在S2亚型中发生频率更高(73%),并且与细胞周期相关的CNA共存,包括9p21.3缺失和11q13.3扩增(图3i-j)。11q23.3缺失的反式效应分析显示,其与细胞周期通路呈正相关,与免疫反应呈负相关(图3k)。11q23.3缺失降低了T细胞和B细胞基因(CD3D、CD3E、CD3G、POU2AF1和IL18)的表达(图3l)。相应地,S2亚型表现出T细胞、B细胞、CD8+ T细胞和自然杀伤细胞浸润减少,同时主要组织相容性复合体(MHC)I类分子(HLA-A/B/C、β2M)和白细胞介素-18(IL-18)的表达也降低(图3l-m)。尽管MHC缺失通常会解除对自然杀伤细胞的抑制,但在S2亚型中,自然杀伤细胞的浸润和受体水平却出人意料地低。这可能是由于11q23.3缺失导致 IL-18 下调,从而损害自然杀伤细胞的募集。因此,11q23.3缺失和CCND1 / MDM2扩增共同驱动 S2 肿瘤中的细胞周期激活和免疫沙漠表型。
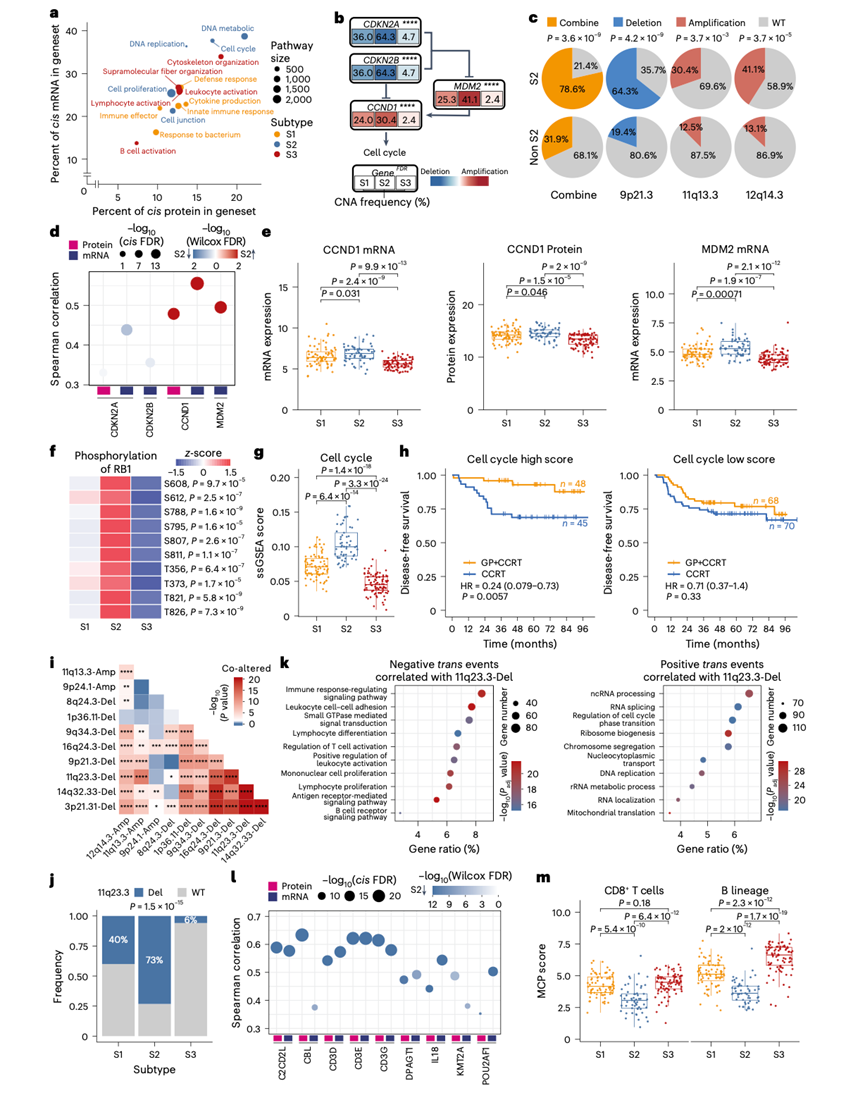
图3. 细胞周期通路的激活驱动 S2 亚型 NPC 患者的 GP-IC 反应。
(a) 按通路划分的顺式mRNA和顺式蛋白比例散点图。(b-c) 细胞周期基因在不同亚型和S2与非S2样本中的拷贝数变异(CNA)频率。(d) 细胞周期基因的顺式效应(CNA-mRNA-蛋白相关性)。(e) 细胞周期基因的表达。(f) RB1磷酸化位点丰度(z评分)的热图。(g) 细胞周期通路的ssGSEA评分。(h) 接受GP+CCRT治疗与仅接受CCRT治疗的患者的DFS曲线,按细胞周期评分分层。(i) 复发性癌症基因普查相关CNA的共现情况。(j) 11q23.3缺失的分布。(k) 与11q23.3缺失相关的反式事件的富集分析。(l) 免疫相关基因的顺式效应。(m) 指定细胞类型的微环境细胞群(MCP)评分。
04
IFNγ 优势的 S1 亚型,具有激活的抗肿瘤免疫力
三种亚型的免疫评分存在差异(图2e)。与此一致的是,xCell对TME的分析显示,S1和S3亚型中免疫细胞浸润程度较高,而S2亚型中浸润程度较低(图4a)。尽管浸润水平相似,但S1和S3肿瘤的临床结局却存在显著差异(图2f-g),提示特定免疫细胞类型与预后相关。S1亚型的特征是存在细胞毒性免疫细胞,包括活化的树突状细胞、M1型巨噬细胞和CD8+效应记忆T细胞(图4a)。此外,S1亚型还表现出细胞毒性因子、MHC成分和趋化因子的上调(图4b),表明存在活跃的免疫反应。S1亚型中与IFNγ反应、抗原呈递和CD8+细胞毒性T细胞相关的通路富集进一步证实了这一点(图2b,图4c)。肿瘤细胞对IFNγ的内在反应是CD8+T细胞介导的抗肿瘤活性的关键决定因素,其机制是通过上调STAT1-IRF1-MHC信号通路。他们观察到S1亚型中STAT1和IRF1表达升高(图4d)。因此,S1亚型代表了一个具有IFNγ反应表型的亚群,其特征是抗肿瘤免疫激活和预后良好。
虽然T细胞浸润丰富,但S3肿瘤缺乏细胞毒性免疫细胞浸润和细胞毒性效应细胞表达,却表现出丰富的B细胞浸润(图4a-b),提示其抗肿瘤免疫受损。此外,S3肿瘤还表现出共抑制受体和T细胞耗竭标志物的升高(图4b和扩展数据图6a),其中CD8+ T细胞耗竭特征显著富集(图4c)。肿瘤微环境(TME)模块分析发现S3亚型中存在一个主要的B细胞模块(图4e),该模块主要由记忆B细胞、经历类别转换重组(CSR)的B细胞和幼稚B细胞组成(图4f),突显了这些B细胞亚群的关键调控作用。在GP+CCRT治疗组中,预后不良的NPC表现出B细胞体液免疫通路富集,以及B细胞/浆细胞(PC)相关蛋白(IGHA1/2、IGHG2/3、IGHV1/3/4/6和JCHAIN)的过表达(图4g-h)。值得注意的是,在CCRT组中,这些蛋白均未出现差异表达(图4h),表明它们对GP-IC疗效具有预测价值。纳入交互项的Cox回归模型显示,IgA +浆细胞分泌的免疫球蛋白A(IgA)同种型(IGHA1/2)在S3亚型中显著过表达,且与GP+CCRT治疗的无病生存期(DFS)或总生存期(OS)获益降低相关(图4i-l)。IgA是黏膜表面最丰富的免疫球蛋白;尽管非黏膜器官中的病理性 IgA 会损害 T 细胞依赖性抗肿瘤免疫,但其在鼻咽癌等黏膜肿瘤中的作用仍不清楚。
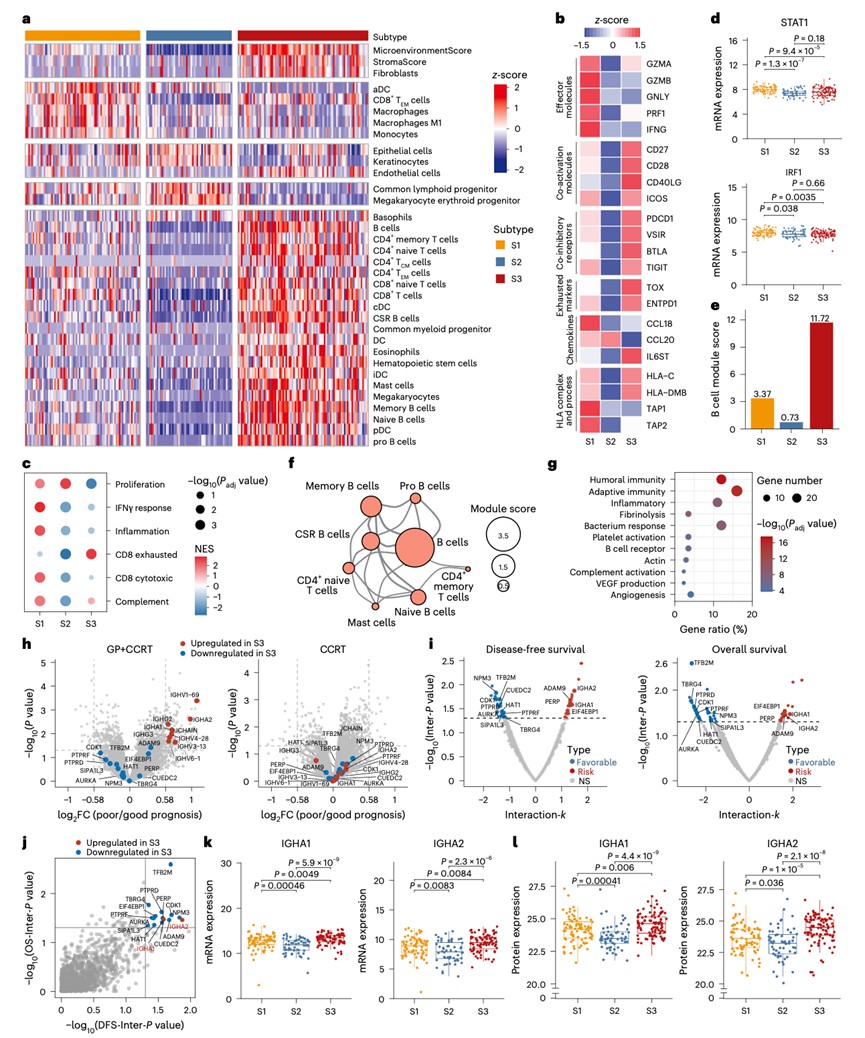
图4. 亚型特异性微环境分析确定 IgA 为 S3 NPC 的治疗生物标志物。
(a) 各亚型中 xCell 衍生细胞特征的热图。(b) 各亚型中免疫效应分子亚群的标准化 mRNA 表达。(c) (GSEA)富集评分的散点图。(d) STAT1(上图)和 IRF1(下图)mRNA 的表达水平。(e) 各亚型中 B 细胞模块评分。(f) S3 亚型的肿瘤微环境(TME)图。(g) 差异表达蛋白 (DEP) 的富集分析。(h) 按治疗组分层,比较预后不良和预后良好的患者的差异表达基因(DEG)。(i) 火山图显示 S3亚型中DEP与治疗组交互作用的DFS和OS的交互P 值和k值。(j) S3亚型中DEP的DFS和OS交互P值。(k-l) IGHA1和IGHA2在不同亚型中的mRNA和蛋白表达水平。
B细胞通过类别转换重组(CSR)产生IgA,该过程需要重链恒定区α基因(Cα)的转录和活化诱导胞苷脱氨酶(AICDA)的表达。S3肿瘤表现出显著高水平的AICDA表达(图5a),且IgA水平与CSR B细胞浸润显著相关(图5b),表明IgA CSR活跃。S3肿瘤中T细胞依赖性(CD40LG)和T细胞非依赖性(TNFRSF13B、TNFRSF13C和TNFSF13B)CSR通路均上调(图5a)。抑制性NF-κB调节因子(IKKA/CHUK和NFKBIA)表达下调,而正向调节因子(NFKB1和RELA)表达上调(图5a)。因此,在S3亚型中, AICDA通过T细胞依赖性和T细胞非依赖性的NF-κB通路通过转录激活,从而促进类别转换重组(CSR)。IgA CSR也需要转化生长因子-β(TGFβ)来启动Cα转录。S3肿瘤表现出TGFβ表达增加,且与IGHA1/2水平密切相关(图5a-d),同时TGFβ受体(TGFβR2和ACVRL1)也上调(图5a)。虽然活性SMAD调节因子并未升高,但SMAD7和SMURF(SMAD泛素化调节因子)的抑制因子表达下调(图5a),这可能维持了TGFβ信号对IGHA转录的激活。总之,AICDA和IGHA均发生转录激活,从而促进S3肿瘤中的IgA CSR。鉴于 S3 肿瘤的 CD8+ T 细胞耗竭表型(图4c和图5e),他们发现 IgA 表达(IGHA1/2)与耗竭的 CD8+ T 细胞评分之间存在正相关性(图5f),表明 IgA CSR 可能导致 S3 肿瘤中 CD8+ T 细胞耗竭和 GP 化疗耐药性。
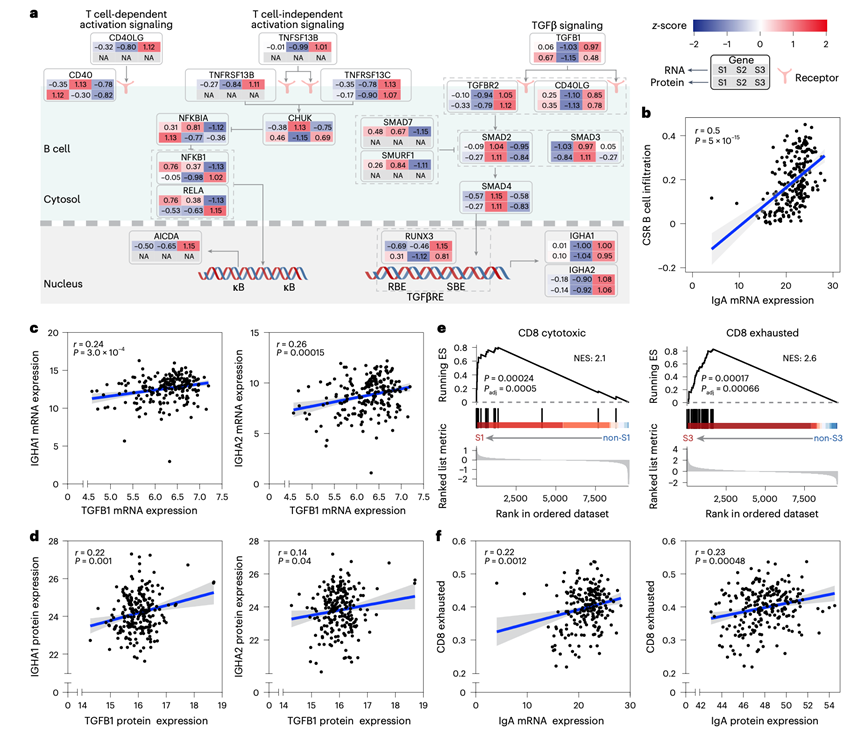
图5. IgA 类别转换与 S3 NPC 中的免疫功能障碍相关。
(a) 示意图展示了三种亚型中IgA类别转换重组(CSR)关键调控因子的mRNA和蛋白表达情况。(b) IgA mRNA表达与CSR B细胞浸润评分之间的相关性。(c-d) TGFB1与IgA在mRNA水平和蛋白水平上的表达相关性。(e) S1和S3蛋白质组亚型置换检验中CD8+ T细胞毒性和耗竭基因集的富集分析。(f) IgA蛋白和mRNA表达与CD8+ T细胞耗竭评分之间的相关性。
05
IgA表达预示GP+CCRT治疗的不良预后
接下来,他们评估了IgA对GP联合CCRT与单纯CCRT治疗的预测价值。总IgA表达的第60百分位数显示出最佳的预测性能,优于单个同种型(IGHA1/2),具有最显著的交互作用P 值和最高的风险k 值(图6a-b)。IgA表达低的患者从GP联合CCRT治疗中获得了显著更长的DFS或OS获益,而IgA表达高的患者则未观察到这种获益(图6c-d)。此外,IgA的预测能力优于包括EBV DNA载量、TNM分期、年龄和性别在内的临床病理特征(图6e-f),这凸显了IgA表达是预测GP+CCRT获益的最佳标志物。在多因素分析中,IgA表达仍然是GP+CCRT获益的强独立预测因子(图6e-f),但在CCRT组中则不然,这突显了其在预测GP-IC获益方面的特异性和独立价值。
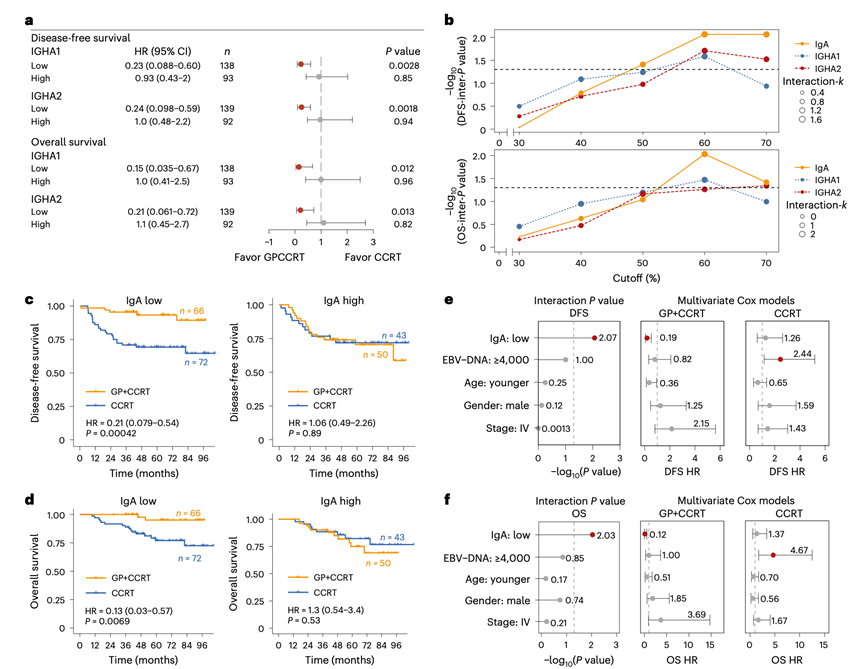
图6. IgA 蛋白表达预测 GP-IC 的临床疗效。
(a) IGHA1 和 IGHA2 蛋白表达与 DFS 和 OS 相关的 HR 值。(b) 计算 IgA 同种型(IGHA1、IGHA2)和总 IgA 表达水平与 DFS(上图)或 OS(下图)的交互作用 P 值和 k 值,以预测 GP + CCRT 与单独 CCRT 治疗的疗效。(c-d) 生存曲线分析。(e-f) 森林图分别显示 GP + CCRT 和 CCRT治疗组中,与治疗组交互作用的临床指标的 DFS 或 OS交互作用P 值(左图)以及多变量 Cox 回归模型的 HR 值(右图)。
06
IgA+ PC-CD8+ T细胞相互作用与GP耐药性相关
他们利用已发表的来自匹配的GP-IC治疗前后NPC样本的scRNA-seq数据进行了研究,将B细胞分为七个聚类(图7a),其中IgA+浆细胞以高表达IGHA1/2为特征(图7b)。化疗后,IgA+浆细胞显著减少,且在治疗有效的患者中减少更为显著(图7c-d),提示IgA+浆细胞的持续存在与化疗耐药相关。Cellchat分析显示,化疗后无应答患者的IgA+浆细胞(PC)与 CD8+ T 细胞之间的通讯增强(图7e-f),主要通过 MIF-CD74 轴介导。此外,CD74 作为 S3 亚型中的关键枢纽,在 S3 肿瘤中显著上调。流式细胞术分析证实,肿瘤内 IgA+浆细胞表现出免疫抑制表型,PD-L1、IL-35 和 MIF 表达升高(图7g-i)。值得注意的是,CD74+CD8+ T 细胞中 PD-1+Tim-3+耗竭细胞群增加(图7j)。重组MIF以剂量依赖的方式诱导外周血单核细胞分离的CD8⁺ T细胞出现耗竭标志物,而MIF/CD74中和抗体可逆转这一过程(图7k)。将CD8⁺ T细胞与鼻咽癌组织来源的IgA⁺浆细胞共培养可显著诱导CD8⁺ T细胞耗竭,而MIF/CD74阻断或IgA⁺浆细胞耗竭可进一步消除这种耗竭(图7l)。这些数据表明,IgA⁺浆细胞通过MIF-CD74轴与CD8⁺ T细胞相互作用,从而驱动T细胞耗竭。
接下来,他们利用多重免疫组化(mIHC)染色验证了“SYSUCC-GP队列”中IgA +浆细胞(PC)与CD8+T细胞的相互作用。S3肿瘤在肿瘤周围区域聚集了大量的IgA+CD138+浆细胞,同时PD-1+CD8+ T细胞浸润也显著增加(图7m-n)。空间分析显示,S3亚型中与PD-1+CD8+ T细胞接触的IgA+浆细胞比例最高(图7o),表明在化疗耐药的S3肿瘤中,这种相互作用增强。与蛋白质组学数据一致,GP-IC显著延长了IgA+浆细胞浸润程度低的患者的生存期,但对IgA+浆细胞浸润程度高的患者则无此作用(图7p-q)。这些发现已在一项独立的多中心III期临床试验队列1(“多中心-GP队列”,队列2)中得到验证。与此一致的是,IgA+浆细胞的增加与PD-1+CD8+ T细胞的频率和空间接触均呈正相关(图7r)。IgA+浆细胞浸润低的患者从GP-IC治疗中显著获益(图7s-t)。Cox回归模型证实IgA+浆细胞与GP-IC治疗的DFS疗效之间存在显著相关性,但与OS疗效的相关性接近显著。总而言之,这些结果表明IgA+浆细胞与PD-1+CD8+T细胞的相互作用与GP化疗耐药相关,并且IgA+浆细胞可能作为鼻咽癌患者GP-IC疗效的预测因子。
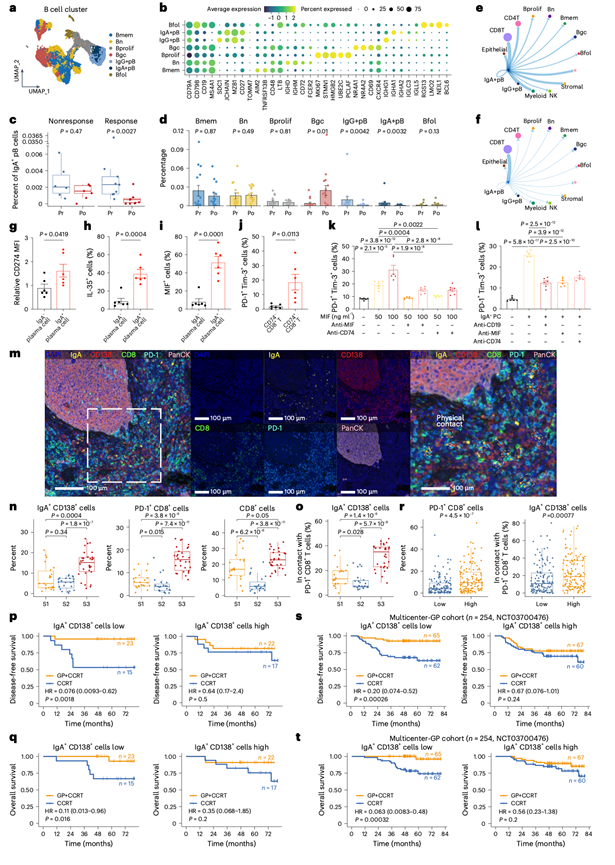
图7. IgA+B 细胞与免疫耗竭和对 GP 化疗的耐药性有关。
(a-d) 均匀流形逼近和投影(UMAP),标记基因热图以及GP化疗前(pr)和化疗后(po)B细胞聚类的组成变化。(e-f) 无应答者中IgA+浆细胞与其他细胞相互作用的数量和权重或强度。(g-j) 平均荧光强度。(k-l) 在MIF处理或IgA +浆细胞共培养下,CD8+ T细胞中PD-1+ Tim-3+的频率。(m) 免疫组化染色结果。(n-o) 各亚型中免疫细胞和物理接触细胞的比例。(p-t)生存分析。
07
高IgA +浆细胞的鼻咽癌患者对化疗免疫疗法的敏感性
最后,他们研究了IgA⁺浆细胞浸润程度高的S3肿瘤的治疗脆弱性。单样本基因集富集分析(ssGSEA)显示,S3肿瘤对PD-1抑制剂治疗的反应性更高(图8a)。S3肿瘤还表现出较高的三级淋巴结构特征评分,这与先前报道的三级淋巴结构与免疫治疗反应改善之间的关联一致。利用空间转录组学(Visium HD,GSE206245),他们分析了接受化疗联合PD-1抑制剂(托瑞普利单抗)治疗的鼻咽癌患者的数据,发现CD8⁺ T细胞是IgA⁺浆细胞的近邻细胞之一(图8b)。在接受额外抗PD-1治疗的患者中,IgA⁺ PC评分显著高于无应答患者(图8c),且与CD8⁺ T细胞耗竭增加和细胞毒性降低相关(图8d)。IgA⁺ PC评分与CD8⁺ T细胞细胞毒性之间的负相关性在应答患者中(图6e–g)强于无应答患者(图8e–i),这与先前PD-1/PD-L1阻断可逆转IgA⁺ PC诱导的CD8⁺ T细胞耗竭的研究结果一致。
他们进一步评估了IgA+ PC是否可以作为预测抗PD-1疗法联合GP+CCRT疗效的生物标志物,这在两项多中心III期临床试验中得到了验证。与“SYSUCC-GP队列”和“多中心-GP队列”的研究结果一致,高水平的IgA+ PC浸润与PD-1+ CD8+ T细胞浸润及其空间邻近性相关(图8j-l)。IgA+浆细胞水平升高的患者从标准治疗(GP+CCRT)联合抗PD-1治疗(信迪利单抗或卡瑞利珠单抗)中获益更多(图8k-m)。总之,高水平的IgA +浆细胞可能作为一种预测性生物标志物,用于识别对GP+CCRT耐药但对联合抗PD-1治疗敏感的鼻咽癌患者。
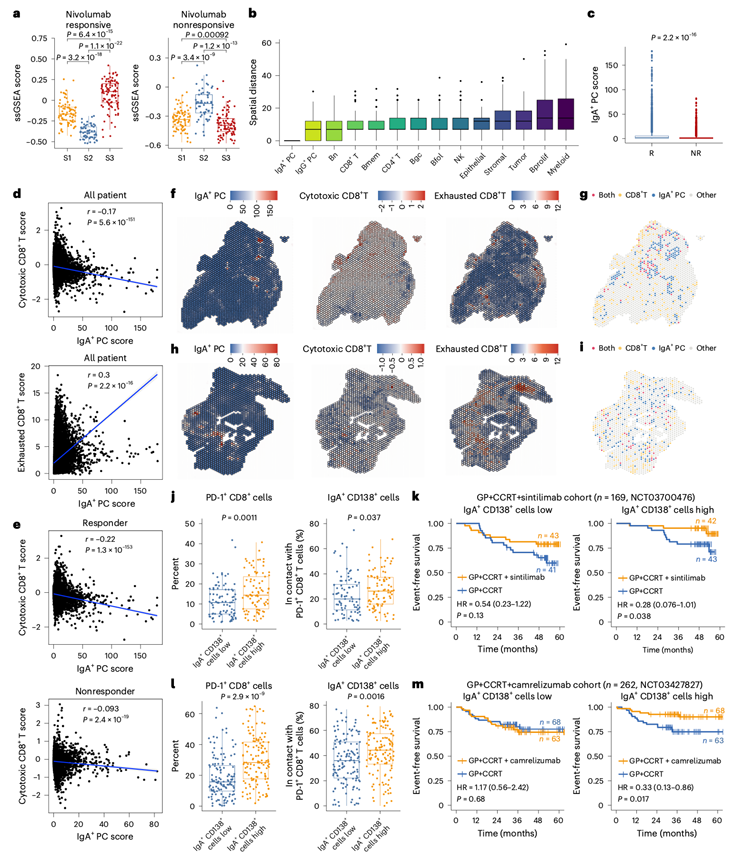
图8. IgA+ CD138+ B 细胞可以预测抗 PD-1 疗法联合 GP+ CCRT 治疗鼻咽癌患者的疗效。
(a) 各亚型中指定通路的ssGSEA评分。(b) IgA+浆细胞与指定细胞类型之间的空间距离。(c) 抗PD-1治疗应答者或无应答者中IgA⁺浆细胞的评分。(d-e) 所有队列患者(d)以及应答者或无应答者中IgA+浆细胞评分与CD8+T细胞细胞毒性和耗竭评分的相关性。(f-i) IgA+浆细胞丰度、CD8+T细胞细胞毒性和耗竭评分的空间分布和共定位情况。(j-m)细胞比例和生存分析。
+ + + + + + + + + + +
结 论
本研究对240例接受GP-IC或CCRT治疗的NPC患者进行了多组学分析,鉴定了三种具有不同治疗敏感性的蛋白质组学亚型。S1亚型主要表现为干扰素-γ反应,且单用CCRT治疗即可获得良好疗效。S2亚型以拷贝数驱动的细胞周期激活为特征,可从GP-IC中获益。免疫耗竭的S3亚型表现出高水平的IgA+浆细胞浸润,对GP-IC耐药,但对PD-1抑制剂治疗有效。单细胞RNA测序证实了无应答者体内IgA+浆细胞与CD8+T细胞之间的相互作用。三项III期临床试验和空间分析的验证表明,高水平的IgA+浆细胞浸润预示着GP-IC耐药,但可从抗PD-1治疗中获益。本研究描绘了GP-IC反应的亚型特异性图谱,并可能为鼻咽癌的个体化治疗提供指导。
+ + + + +



