

 English
English文献解读|Cancer Cell(44.5):弥漫性大B细胞淋巴瘤蛋白基因型的发病机制
✦ +
+
论文ID
原名:Pathogenesis of diffuse large B cell lymphoma proteogenotypes
译名:弥漫性大B细胞淋巴瘤蛋白基因型的发病机制
期刊:Cancer Cell
影响因子:44.5
发表时间:2026.06.04
DOI号:10.1016/j.ccell.2026.05.008
背 景
弥漫性大B细胞淋巴瘤(DLBCL)是一种在遗传和临床上均具有异质性的疾病。尽管约60-70%的DLBCL患者可通过R-CHOP化疗联合免疫疗法治愈,但仍有30-40%的患者会复发或对治疗无效。基因表达谱分析揭示了两种主要的亚型:生发中心B细胞样(GCB)和活化B细胞样(ABC),它们分别与未转化的生发中心B细胞或通过B细胞受体(BCR)活化的B细胞共有基因表达特征。虽然不同细胞起源(COO)亚型之间的突变模式和对免疫化疗的反应存在显著差异,但DLBCL的许多异质性仍未得到充分阐明。例如,10-15%的病例无法通过基因表达谱分析进行分类,并且多项研究强调了同一亚型内不同患者的临床结局存在差异。后续研究建立了包含七个主要遗传亚型的遗传分类,每个亚型都具有特定的基因组改变模式。不同的遗传特征与ABC或GCB基因表达谱相关。例如,BCR亚基CD79B和免疫信号转导衔接蛋白MYD88发生突变的肿瘤(MCD/C5遗传亚型)是ABC型DLBCL中最常见的类型。相比之下,EZH2突变和BCL2易位(EZB/C3)的病例主要表现为GCB表型。重要的是,一些遗传改变与特定的生存通路相关,这些通路可用于治疗。
实验设计
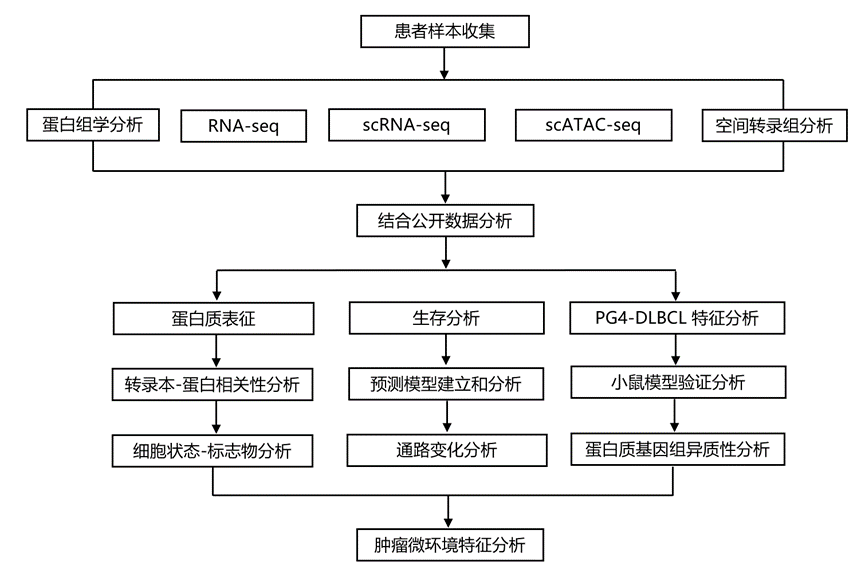
结 果
01

基于多组学的弥漫性大B细胞淋巴瘤蛋白基因型发现与验证
研究团队采用蛋白质组学多平台方法,对来自332例未经治疗的DLBCL患者的福尔马林固定石蜡包埋(FFPE)肿瘤活检样本(发现队列)进行了分析。该队列涵盖了所有COO亚型和临床风险类别,与已发表的接受R-CHOP方案治疗的DLBCL队列相似。他们使用NCI LymphGen和英国HMRN分类器对各个DLBCL基因亚型进行了分类,两者显示出很强的相关性,从而相互验证了先前发表的结果。值得注意的是,由于缺乏拷贝数变异数据,无法识别A53基因亚型。除了这些遗传和基因表达数据外,他们还利用基于串联质谱标签(TMT)的定量质谱技术分析了DLBCL的蛋白质组数据(图1A-B)。获得了高质量的蛋白质组数据集,其中包含6980种蛋白质的定量数据,且未发现与基于TMT的多重分析方法或组织来源相关的显著批次效应,也未发现可能干扰大规模蛋白质组学研究的缺失值模式。
既往研究报道不同肿瘤类型中转录本和蛋白质表达水平的相关性较差,表明转录后/翻译机制对肿瘤病理生理学有显著影响。因此,他们假设整合蛋白质组学方法可能比单纯基因组学方法更具优势,有助于进一步解析DLBCL的生物学和临床异质性。事实上,即使去除可能导致变异性增加和相关性降低的低丰度蛋白质,他们检测到的转录本-蛋白质对的总体相关性仍然较低(图1C)。值得注意的是,定义DLBCL COO亚型的特定转录本-蛋白质对子集显示出高度相关性,表明这些特征较少依赖于转录后/翻译过程(图1D)。
为了整合这些不同的数据集,他们采用了多视图潜在变量建模(MuVI),这是一种概率方法,它基于SignatureDB数据库中反映正常和恶性淋巴细胞生物学特性的基因特征,推断可解释的潜在因子(LF)(图1E)。MuVI能够识别出哪些LF子集驱动了每种实验模式下的患者间差异,并为每位患者分配一个LF活性评分。在推断过程中,MuVI通过移除或添加单个特征来进一步优化SignatureDB特征集。在本研究数据集中,MuVI揭示了140个驱动分子患者间异质性的LF。其中包括11个与COO相关的特征集,COO是已知的变异来源,这些特征集能够反映主要的基因表达亚型,即ABC型和GCB型DLBCL。除了 COO 之外,与 DLBCL 基质浸润和增殖相关的 LF 也导致了患者间的差异,已知这两者都与接受 R-CHOP 方案治疗的 DLBCL 患者的生存率相关。
为了探索其他分子因素对变异的贡献,他们重点关注了127个与COO、恶性细胞增殖或肿瘤免疫/基质含量无关的、能够解释数据变异的LF。然后,使用社群检测方法对这些剩余因子进行了基于图的共识聚类。结果在共识聚类的所有迭代中均稳定地鉴定出了7个DLBCL蛋白基因型(PG)(图1 F-G)。有趣的是,与PG呈正相关的LF在转录组学和蛋白质组学数据集上对变异的解释力不同,这凸显了整合多组学方法的优势(图1 H)。
为了验证这些DLBCL的PG,他们利用转录组分析(RNA-seq)技术和基于质谱的蛋白质组技术进行分析,对包含146例肿瘤的独立DLBCL队列(验证队列)进行了表征。然后,利用发现队列的转录组和蛋白质组数据集,训练了一个梯度提升树学习分类器,以多类别分配方法预测验证队列中的PG。使用一个预留的测试集(来自发现队列)评估了所得的分类器模型,结果显示其性能指标优异(图1 I)。观察到的PG频率与发现队列中的频率一致(图1 J)。为了进一步验证分配结果,使用MuVI方法整合了验证队列的转录组和蛋白质组数据集。接下来,将验证队列多模态表达数据的潜在表示投影到发现队列的多模态潜在表示中(图1K),以量化预测的PG之间的相似性。结果显示,在发现队列和验证队列中识别出的各个PG之间具有高度的分子相似性(图1L),即分配到同一PG的肿瘤在潜在空间中映射紧密,且与它们所属的队列无关。此外,两个队列中PG特异性分子特征的相关性也很好,验证了PG分类。
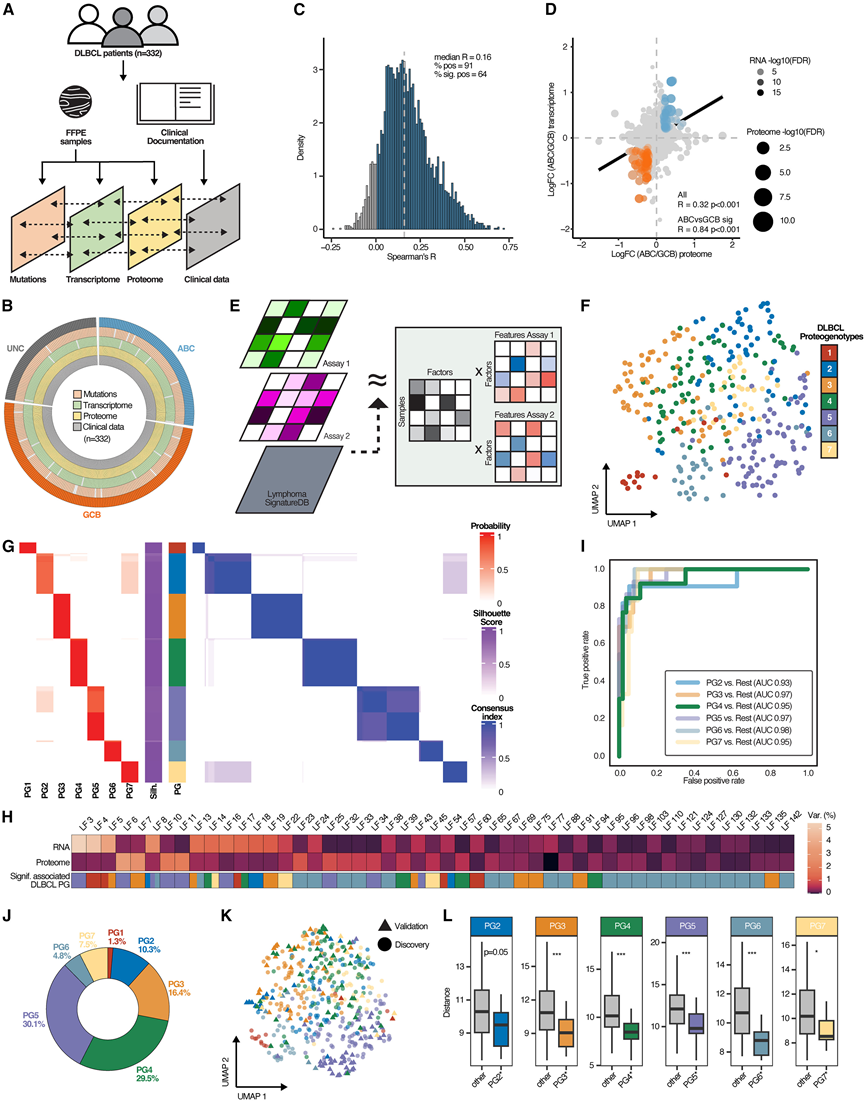
图1. 通过整合蛋白质基因组学鉴定和验证DLBCL蛋白质基因型。
(A) FFPE DLBCL 组织表征的整合蛋白质基因组学方法。(B) 所有数据层中每个样本的数据完整性。(C) 所有检测到的mRNA/蛋白质对的转录组与蛋白质组表达相关性。(D) ABC 型与 GCB 型弥漫性大B细胞淋巴瘤 (DLBCL) 中差异表达的 mRNA(y 轴)和蛋白质(x 轴)的变化。(E) 计算多组学整合方法(MuVI)。(F) 细胞起源(COO)校正的多组学潜在空间的UMAP。(G) 对 COO 状态进行校正后,基于图的共识聚类推断潜在空间的结果指标。(H) 与蛋白质基因型显著相关的潜在因子所解释的方差。(I) 使用在发现队列上训练的 XGBOOST 模型进行预测的接收者操作特征(ROC)。(J) 验证队列中预测的DLBCL蛋白基因型的分布。(K) 发现队列和验证队列联合潜在空间的UMAP图。(L) PG预测的验证队列样本到所有发现队列样本质心的欧氏距离,指示了联合潜在空间内的蛋白质基因型。
02
DLBCL蛋白基因型的独特分子和细胞特征
为了进一步探究PG的特征,他们研究了它们与先前报道的DLBCL分子模式之间的关系。这七个PG在COO和基因亚型方面存在显著差异。PG1和PG7几乎全部由GCB病例组成。PG2富集了EZB亚型的GCB肿瘤。相比之下,PG6富集了未分类(UNC)病例。虽然PG4肿瘤主要为ABC型弥漫大B细胞淋巴瘤,但该PG组也包含相当数量的GCB型和UNC型亚型(图2A)。ABC-PG4病例显著富集MCD(HMRN MYD88)基因亚型(图2A-B),但并非仅限于此;而GCB-PG4肿瘤则富集双重打击特征(DHITSig)阳性,高于所有其他(非PG4)GCB型(图2A-B)。值得注意的是,14例DLBCL病例中仅有2例归类为PG4。既往研究表明,仅约50%的DHITSig+ GCB病例同时经FISH证实为携带MYC/BCL2易位的双重打击肿瘤(图2A)。因此,PG4中DHITSig+肿瘤富集,但缺乏MYC重排。总体而言,各PG组间IPI评分分布无显著差异(图2A-B)。
最近,基于转录组数据的LE框架对DLBCL进行了表征,揭示了反映B细胞分化连续体的细胞状态,其中一些状态具有预后意义。PG5和PG6均富集B细胞状态2,其特征是表达前体记忆B细胞标志物。PG3和PG7则过度表达B细胞状态1,该状态代表生发中心(GC)相关的B细胞表型,预后良好,这与这些PG中GCB肿瘤的高发生率相符(图2A-B)。相反,ABC-PG4富集B细胞状态5,具有GC暗区和浆母细胞特异性基因和蛋白表达特征(图2A-B)。
为了进一步研究与PG相关的分子特征,他们利用SignatureDB进行了基因集变异分析(GSVA),比较了每个PG与其他所有PG(图2C)的差异基因和蛋白质表达集。在PG3和PG7中,GSVA评分显示与GC和静止B细胞以及GCB型DLBCL相关的转录组和蛋白质组特征较高,这与这些PG中GCB病例和B细胞状态1的富集相一致(图2C)。PG5显示出基质和间质特征集的富集(图2A-C)。相比之下,PG4的特征是转录组和蛋白质组特征反映了GC中心母细胞和浆细胞样B细胞状态、MYC驱动的转录、增殖和IRF4信号传导(图2C)。值得注意的是,在PG4组中,MCD和DHITSig+ GCB肿瘤均显著富集(图2A-B)。如上所述,PG4组中核糖体特征集的富集主要体现在蛋白质组水平。反映炎症表型的特征集,包括干扰素信号通路、CD4+和CD8+ T细胞以及髓单核细胞生物学特征,在PG6组中富集,而在PG4组中则显著减少(图2C)。
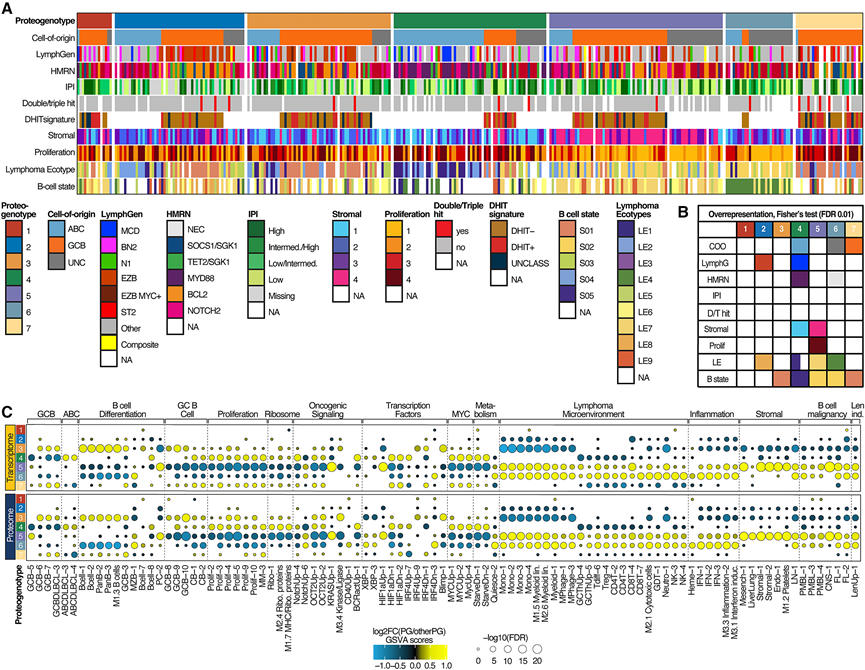
图2. DLBCL蛋白基因型的表型特征。
(A) 已鉴定的每种 DLBCL 蛋白基因型(PG)的疾病相关和临床元数据。(B) 对每种蛋白质基因型的元特征进行Fisher检验,以分析其过度表达情况。(C) 与所有其他蛋白质基因型相比,每个蛋白质基因型的差异GSVA得分。
03
DLBCL蛋白基因型具有预后价值
为了探究各个PG是否具有预后意义,他们比较了它们在R-CHOP治疗后的临床结局。值得注意的是,观察到不同PG组的总生存期(OS)和无进展生存期(PFS)存在显著差异(图3A-D)。尤其值得注意的是,PG4的OS和PFS显著低于发现队列中的所有其他PG组(图3B-D)。在校正多重假设检验后,这种统计学差异仍然存在。在包含ABC基因表达状态和IPI风险因素作为协变量的多因素分析中,PG4已确定为OS较差的独立危险因素(图3E)。此外,PG4较差的OS也独立于先前发表的转录组风险组A7相关的不良生存期,该组的特征是ABC肿瘤富集和免疫细胞浸润低。相比之下,富含免疫细胞的 PG6 与其他所有病例相比,生存趋势更好,其 OS 和 PFS 明显优于 PG4。
为了将此分析扩展到其他缺乏蛋白质组学测量的DLBCL队列,基于DLBCL发现队列的转录组数据训练了一个梯度提升树学习模型,并仅使用RNA表达数据预测了PG。在训练过程中,该分类器在预留测试集上评估时表现出优异的性能指标。该分类器工具可在https://dlbcl-pg-classifier.uni-frankfurt.de/公开使用。随后,他们将该PG-转录组分类器应用于独立的DLBCL队列(NCI和BCC队列)(图3 F)。结果表明,这些队列的PG在细胞起源状态和淋巴基因分型方面与发现队列相似(图S3 F)。此外,预测的PG4病例预后较差,与发现队列相似(图3G-H),且与COO(ABC)、MCD基因亚型或高IPI无关。值得注意的是,在所有三个研究队列中,PG4在最初12个月内的PFS事件发生率均高于其他所有PG。综上所述,通过转录组学或多组学方法鉴定的DLBCL PG在R-CHOP治疗中具有不同的临床结局,其中PG4的生存期最差。
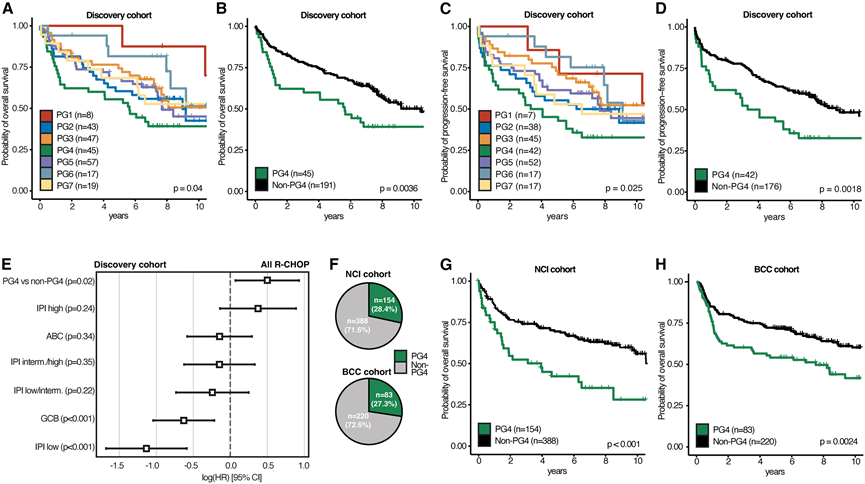
图3. DLBCL蛋白基因型的预后意义。
(A) 按 DLBCL 蛋白基因型分层的总生存期(OS)估计值。(B) 按 PG4 与所有其他 DLBCL 蛋白基因型分层的 OS 估计值。(C) 按 DLBCL 蛋白基因型分层的无进展生存期(PFS)估计值。(D) 按PG4与其他所有DLBCL蛋白基因型分层的PFS估计值。(E) 采用多变量Cox回归分析,比较PG4与其他DLBCL蛋白基因型,并将IPI评分和细胞起源状态作为协变量。(F) NCI 和 BCC 队列中 PG4 预测病例的比例。(G) NCI队列中预测的PG4与所有其他蛋白质基因型的OS估计值。(H) 预测的PG4与BCC队列中所有其他蛋白基因型的OS估计值。
04
PG4-DLBCL 的独特生物学特性
鉴于PG4预后不良,他们接下来对其潜在的生物学机制进行了表征。鉴定了五个基于通路的LF,它们解释了PG4与其他所有PG之间的差异(图4A-B)。定义这些LF的特征与SignatureDB中先前的特征集在多模态建模后显示出显著重叠(图4B)。重要的是,只有两个PG4驱动LF在转录组层面上具有活性,而所有五个LF均解释了蛋白质组层面的差异,这凸显了它们捕捉相关疾病特征的额外能力(图4B)。接下来,通过评估MuVI权重,他们探索了定义PG4特异性LF的分子特征(图4B-C)。结果表明,这五个蛋白质组LF展现出一组复杂的特征。所有五个LF均揭示了与B细胞明暗区生物学相关的特征,这些特征并非相应稀疏先验的一部分,同时还富集了与核糖体生物学和蛋白质翻译相关的通路。这些特征均未在转录组水平上捕获(图4C)。
为了进一步揭示PG4的分子和细胞特征,他们比较了所有COO亚组中PG4和非PG4的转录组表达数据所对应的SignatureDB平均特征评分。由于单个基因集中的转录组信息并不一定能转化为相似的蛋白质组特征集(因为它们的相关性总体较低),他们将SignatureDB中涵盖的表型转录组信息转化为信息丰富的蛋白质集,以便对蛋白质组数据进行更深入的探索。在此过程中,他们发现了几个相关的模式。首先,在转录组和蛋白质组数据集(即“数据层”)中,PG4中GC T滤泡辅助细胞和滤泡树突状细胞的特征均显著减少(图4D)。其次,无论细胞来源如何,PG4 均富集了源自CCR6+记忆 B 前体细胞(Bcell-4)和(前)浆母细胞(Blimp-2、Bcell-8)以及生发中心暗区细胞(CB-1、GCB-10)的特征(图 4E)。GC暗区生物学此前已证实与 GCB 型DLBCL以及该亚型小鼠模型中的MCD 型 ABC 型 DLBCL 相关。第三,与非PG4病例相比,ABC-PG4和GCB-PG4病例中BCR信号通路特征(BCRactUp-1)和MYC活性的表达均显著升高,提示PG4基于共同的致病特征将来自不同COO背景的DLBCL联系起来(图4F)。
除了MYC相关基因集外,PG4还高表达由OCT2转录激活的基因和蛋白(OCT2Up-1),OCT2是一种转录因子(TF),是生发中心B细胞浆细胞分化所必需的(图4F)。一个值得注意的OCT2直接靶基因是XBP1,它是一种TF,能够建立浆细胞的分泌表型,并协调核糖体生物合成和总蛋白合成。有趣的是,XBP1特征以及核糖体亚基和蛋白质翻译特征对PG4特异性蛋白质组学LF有显著贡献(图4C),并且核糖体蛋白质组学特征在所有COO亚型的PG4中均显著上调(图4G)。PG4核心蛋白质组包括核糖体结合蛋白、mRNA相互作用蛋白和与多聚核糖体相关的蛋白。最后,在COO亚型中,PG4与BCR、PI3激酶(PI3KUp-1)和mTOR信号通路(RapaDn-2)的特征相关(图4 F)。重要的是,PG4 定义的分子特征在发现队列和验证队列之间具有很强的相关性。
接下来,他们探讨了特定致癌突变是否在各个PG中富集(图4H)。结果显示,PG1中IRF8突变富集,PG2中STAT6突变富集,PG5中EZH2和TET2突变富集,PG7中EZH2、BCL2、TNFRSF14、REL和SGK1突变富集。对于PG4,ABC-PG4病例显示出与MCD亚型相关的突变富集(图4I),包括TBL1XR1和BTG1突变,已知这两个基因在淋巴瘤发生中起重要作用(图4H)。作为SMRT/HDAC3复合物的一部分,突变型TBL1XR1会使该蛋白复合物的相互作用偏向于BACH2,而减弱其与BCL6的相互作用,从而改变生发中心B细胞的特性并促进其分化。BTG1体细胞突变会产生一种超强竞争性的恶性B细胞表型,并在淋巴瘤小鼠模型中诱导MYC相关的生物合成特征,包括相关核糖核苷酸和核糖体相关蛋白的上调,这与上述PG4的特征一致。
总之,PG4 显著富集 ABC MCD (MYD88) 肿瘤,尤其是携带BTG1和TBL1XR1突变的肿瘤(图 4I-J),以及 DHITSig+ GCB 肿瘤(图4I-J)。尽管 PG4 跨越了 COO 边界,但其统一特征包括 GCB 暗区表型和免疫细胞耗竭的肿瘤微环境 (TME)。PG4 的其他标志性特征包括 MYC 活性增强和蛋白质翻译机制增强。
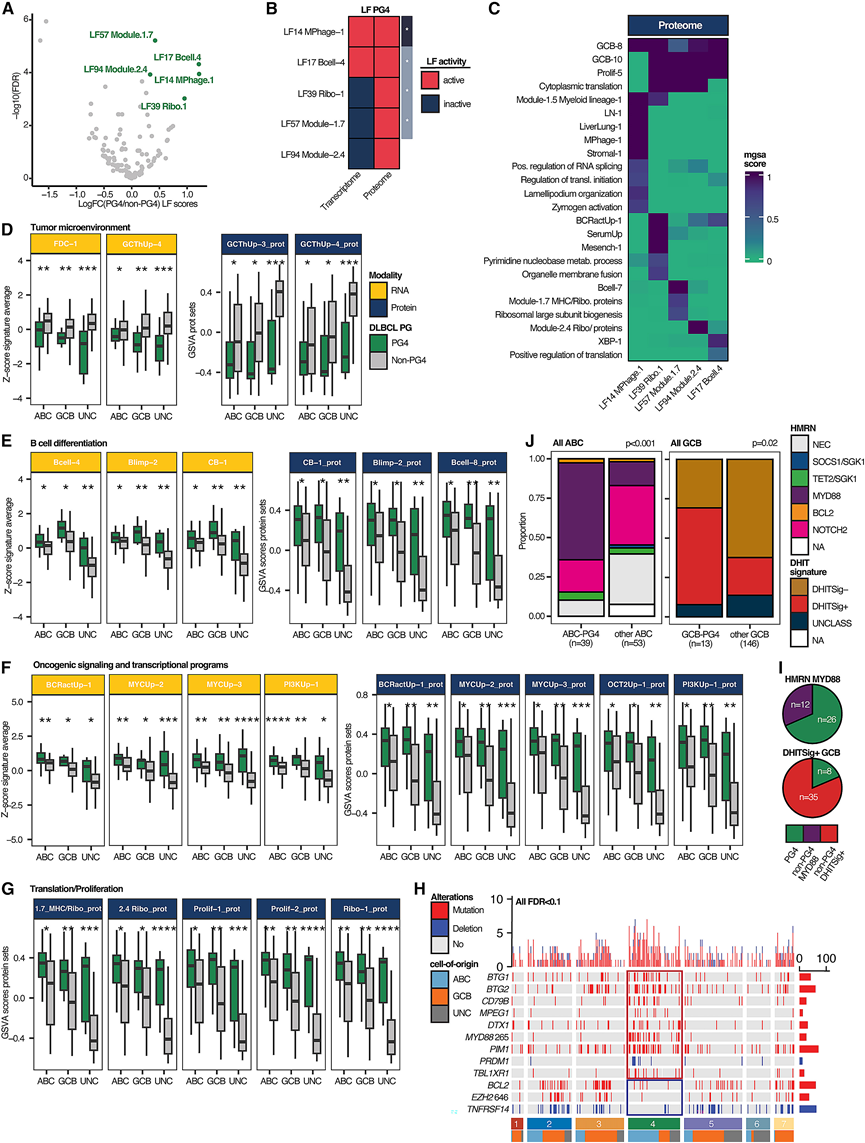
图4. PG4-DLBCL 的特征是免疫抑制性肿瘤微环境和细胞内在通路失调。
(A) 对LF与DLBCL的PG4状态进行显著性关联的差异性检验。(B) PG4驱动因子活性热图。(C) 贝叶斯调节富集分析与基于模型的基因集分析。(D) 根据细胞起源状态,PG4 与所有其他非 PG4 病例之间,基于转录组(左图)的与 TME 相关的显著差异平均特征评分,以及从 SignatureDB 对应的与 TME 相关的表型蛋白质集(右图)得出的显著差异 GSVA 评分。(E) 与 (D) 类似,这里是与 B 细胞分化相关的特征集。(F) 与 (D) 和 (E) 类似,用于与致癌信号和转录因子活性相关的特征集。(G) 基于 SignatureDB 对应表型蛋白集(蓝色)的 GSVA 评分,在 PG4 和其他所有非 PG4 病例中,与蛋白翻译和增殖相关的特征集存在显著差异。(H) Oncoprint 突出显示了 PG4-DLBCL 与其他所有突变相比显著过度(红色框)或不足(蓝色框)的突变。(I) 所有 HMRN MYD88 病例中的 PG4 病例数(上图)和所有 DHITSig+ 病例中的 PG4 病例数(下图)。(J) PG4 ABC 和非 PG4 ABC 病例中 HMRN 病例的相对分布(左图)和 PG4 GCB 和非 PG4 GCB 病例中 DHIT 特征病例的相对分布(右图)。
05
BTG1突变和MYC参与PG4-DLBCL的发病机制
BTG1突变已确定为ABC型和GCB型PG4肿瘤的遗传标志。然而,它们对PG4发病机制的影响尚不清楚。为了研究这一点,他们对经VavP-Bcl2基因改造并敲入Btg1 Q36H突变的小鼠进行了淋巴瘤实验评估(图5A)。小鼠和人类的BTG1蛋白序列相同,且BTG1 Q36H突变在人类DLBCL中高度复发。在该异位表达BCL2的小鼠淋巴瘤模型中,B细胞中BTG1 Q36H的条件性表达显著加速了淋巴瘤的发生,导致淋巴组织结构破坏、侵袭非淋巴组织,并降低了小鼠的生存率,而仅表达Bcl2转基因的小鼠则无此现象。对这些小鼠淋巴瘤的转录组分析显示,与野生型BTG1小鼠相比,BTG1 Q36H突变小鼠的MYC和PG4核心特征评分显著升高(图5B)。他们还通过在人GCB细胞系(SUDHL-4)中异位表达Btg1 Q36H突变体来模拟其功能,并观察到与过表达野生型BTG1的细胞相比,MYC和PG4特征的表达也出现了类似的升高(图5C)。使用ChEA3流程进行的转录因子富集分析也显示,与BTG1野生型细胞相比, BTG1 Q36H突变细胞中MYC和TCF3调控的基因富集(图5D)。最后,通过免疫组化(IHC)检测了BTG1 Q36H小鼠淋巴瘤的TME,观察到T细胞区室异常,表现为CD8+ T细胞减少和剩余CD4+ T细胞分散(图5E)。因此,BTG1 Q36H小鼠模型重现了PG4表型的关键特征,包括定义恶性细胞的转录因子网络以及相关的TME改变。
他们采用正交方法,通过将一系列人类DLBCL细胞系归类为不同的PG,进一步验证了MYC活性失调作为PG4的标志。他们对53株DLBCL细胞系进行了RNA-seq和蛋白质组学分析,并使用建立的多模态PG分类器进行PG分类(图5F)。与归类为其他PG的细胞系相比,PG4样细胞系模型具有显著更高的PG4核心蛋白组表达和MYC靶蛋白表达(图5G)。此外,他们观察到PG4样细胞系模型的转录组谱与发现队列中原发性PG4肿瘤的转录组谱之间存在高度相关性。
为了从功能上评估MYC在PG4中的活性,他们选择了6株PG4样细胞系和8株非PG4样细胞系,并分别用MYC/MAX抑制剂或DMSO处理这些细胞系。24小时后,通过质谱分析进行了全蛋白质表达谱分析。差异表达蛋白的富集分析显示,在PG4样细胞系中,MYC靶基因特征(MYCUp-4)和PG4核心蛋白质组特征显著下调,而在非PG4样细胞系中则未观察到此现象(图5H)。因此,MYC活性是PG4的标志性特征,而BTG1突变(以及其他潜在的基因改变)的获得进一步强化了这一特征。
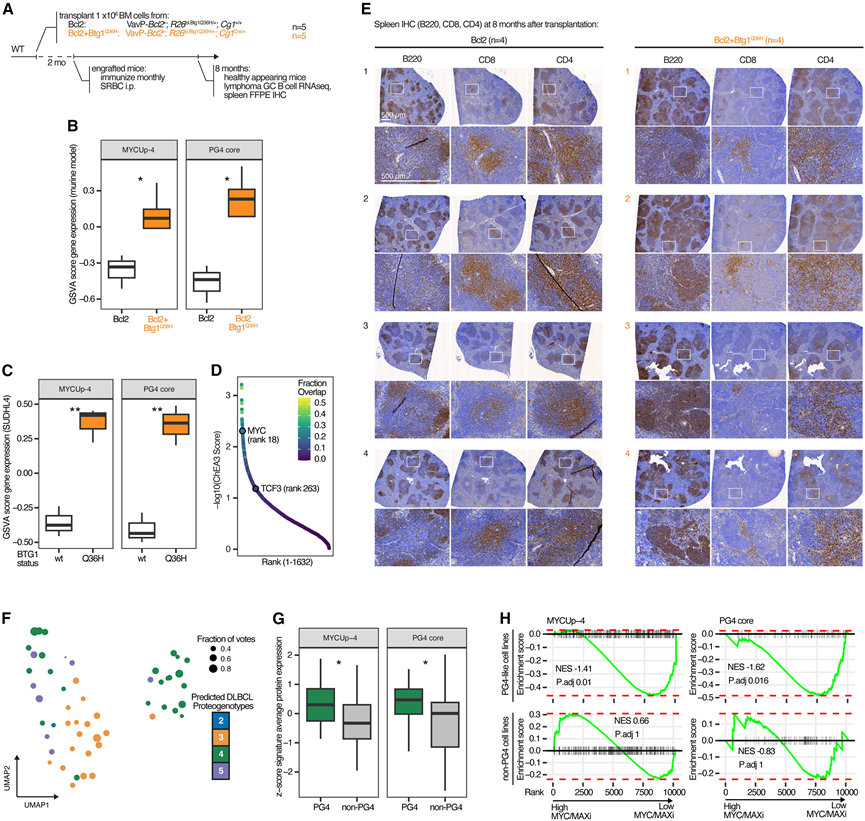
图5. 高 MYC 活性是 PG4-DLBCL 的标志。
(A)实验设计。(B-C) GSVA 分析。(D) 对转录因子 (TF) 进行排序。(E) 免疫组织化学(IHC)染色。(F) 基于 53 个 DLBCL 细胞系模型的转录组和蛋白质组表达数据的多组学潜在空间 UMAP。(G) 与 MYC 靶标(MYCUp-4)和 PG4-DLBCL 核心蛋白相关的特征集的特征平均得分。(H) 与 MYC 靶标(MYCUp-4)和 PG4-DLBCL 核心蛋白相关的特征集的特征平均得分。
06
单细胞分辨率下解析的PG4-DLBCL转录特征
受蛋白质组学分析结果的启发,他们利用来自蛋白质组学验证队列中43例DLBCL患者活检组织的单细胞转录组分析(scRNA-seq)和单细胞ATAC测序(scATAC-seq)数据,探索了PG4的生物学机制(图6A)。该样本集中的COO亚型分布与DLBCL总体分布具有代表性。他们使用多组学分类器将肿瘤分类为PG,并在43例肿瘤中鉴定出13例为PG4。使用加权最近邻(WNN)分析方法整合了scRNA-seq和scATAC-seq数据,该方法基于降维后的数据表示。通过UMAP可视化推断的基于WNN图的表示,鉴定出了TME区室,其中包括CD4+和CD8+ T细胞、髓系细胞、基质细胞以及正常和恶性B细胞(图6B)。值得注意的是,DLBCL的PG中恶性细胞、基质细胞和免疫细胞的比例各不相同。与整体蛋白质组学数据一致,PG6的TME最为丰富,含有大量的T细胞和其他非恶性TME细胞类型,而PG4肿瘤则以恶性B细胞为主,TME较为稀疏。为了更详细地比较不同PG的细胞组成,他们利用k近邻图推断了具有相似基因表达的细胞邻域,并鉴定出特定PG细胞显著富集的邻域。他们首先关注PG4病例中细胞显著富集的恶性B细胞邻域(图6C)。在每个COO亚组中,鉴定出与非PG4恶性B细胞邻域相比,在这些PG4优势恶性B细胞邻域中丰度富集的转录本。特征分析揭示了PG4中富集的生物学主题,这些主题与以下方面相关:1)MYC靶基因(MYCUp-1、MycUp-4);2)核糖体功能(模块1.7、模块2.4、Ribo-1);以及3)GC B细胞亚群,包括暗区细胞(GCB-10)和中间区细胞(GCB-9)(图6D-E)。
接下来,他们研究了PG4是否具有特定的染色质可及性模式。事实上,通过将基因表达估计与scATAC-seq染色质可及性数据整合,鉴定出一组PG4特异性的染色质特征,这些特征分别表明GCB-PG4和ABC-PG4中TCF3和TCF4转录水平的过度激活。TCF3是一种驱动生发中心暗区表型的转录因子,并且在伯基特淋巴瘤中具有特征性的活性。在PG4恶性B细胞邻近区域的可及染色质峰中,TCF3和ID3相关的结合基序显著富集(图6F)。值得注意的是,TF富集分析预测MYC、TCF3和TCF4负责PG4特异性蛋白特征的表达,从而表明这些TF参与了PG4的发病机制(图S6 D)。他们发现PG4恶性B细胞中ATAC-seq开放染色质峰与染色质免疫沉淀测序(ChIP-seq)确定的已知TCF3或TCF4结合位点存在显著重叠,且18q扩增无显著差异。
此外,ABC-PG4 中 IRF4 的靶基因显著富集。IRF4 是一种转录因子,在 ABC DLBCL 中由 BCR 依赖的 NF-κB 信号通路诱导表达。IRF4通过分别与 BATF 或 SPIB 的 AICE 或 EICE 结合基序配对,促进靶基因表达。在 PG4 中,观察到 IRF4 的 EICE 和 AICE 基序在开放染色质峰中高度富集(图 6F),这通过转录因子足迹分析得到证实。
在对PG4-DLBCL的淋巴瘤细胞固有特征进行表征后,他们接下来探索了这些PG4特征是否能在DLBCL肿瘤内的基因亚克隆中检测到。为此,利用了102例DLBCL病例的单细胞测序数据,这些病例的肿瘤亚克隆是通过DNA拷贝数增减模式识别的。为了鉴定PG4样亚克隆,他们对每个亚克隆中所有恶性B细胞的单细胞转录组数据进行了伪批量聚合,并将其输入到转录组分类器中。他们以高置信度鉴定出243个恶性B细胞亚克隆中的89个为PG4样亚克隆。重要的是,相当数量的肿瘤至少含有一个PG4样亚克隆,这些亚克隆通常占肿瘤中恶性B细胞的比例不到20%(图6G-H)。LymphGen PG4 样亚克隆和 PG4 肿瘤的基因亚型分配与发现队列相似。此外,PG4 样亚克隆和 PG4 肿瘤的基因表达谱均表明 BCR-PI3K 通路活性、MYC 靶基因富集以及 GC 暗区 B 细胞表型。
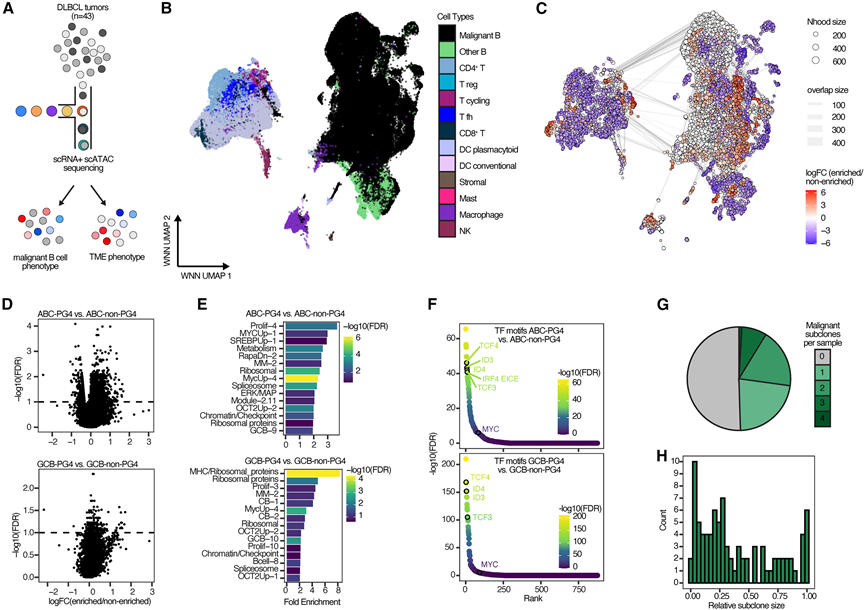
图6. PG4-DLBCL 中的单细胞基因表达和染色质图谱。
(A) PG4-DLBCL 中的单细胞基因表达和染色质图谱。(B) 验证队列中 43 名 DLBCL 患者的 197,860 个单细胞的 RNA 表达和 ATAC 测序数据的加权最近邻整合维度的 UMAP。(C) 基于图的推断单细胞邻域可视化。(D) 在 ABC(上图)或 GCB(下图)中,PG4 过度表达的恶性 B 细胞邻域与非 PG4 恶性 B 细胞邻域的标记基因检测。(E) 使用 SignatureDB 作为特征集集合,对 PG4 恶性 B 细胞邻域标记进行富集分析的超几何检验,分别在 ABC(上图)或 GCB(下图)中进行。(F) 与ABC病例(上图)或GCB病例(下图)中所有其他非PG4恶性B细胞邻近区域相比,PG4恶性B细胞邻近区域中转录因子(TF)基序的富集情况。(G) 单细胞数据集中每个样本的PG4预测恶性B细胞亚克隆数。
07
DLBCL蛋白基因型中免疫细胞和基质细胞分布的差异
蛋白质组学分析除了揭示淋巴瘤细胞固有特征的失调外,还发现不同PG的肿瘤微环境(TME)细胞组成存在差异(图2)。对发现队列的多组学数据集的进一步探索表明,PG6中淋巴瘤生态型4(LE4)过度表达,该生态型以CD4+、CD8 +和调节性T细胞以及单核细胞的高丰度为特征。PG6的炎症性TME特征通过炎症性TME特征的高GSVA评分得到独立验证。相反,PG5中LE7富集,代表以树突状细胞和滤泡辅助性T细胞为主的TME,并且也显示出较高的间质、基质和生发中心样TME特征评分。这与CIBERSORT.x推断出的成纤维细胞、内皮细胞和CD4+ T细胞相对丰度较高相符。12相反,PG4中CD4+和CD8+ T细胞数量减少,且表达的免疫细胞耗竭的肿瘤微环境(TME)特征也降低。13为了进一步在单细胞水平上探索TME,他们分析了PG富集的单细胞区域中免疫细胞和基质细胞类型的相对丰度。结果显示,PG4在CD4 +和CD8+ T细胞区域中显著减少,尤其是在CD4+ T滤泡辅助细胞区域中,这证实了整体蛋白质组学数据(图7 A)。相比之下,PG2 肿瘤在 T 调节细胞和 CD8+ T 细胞群的各种 T 细胞邻域中显著富集(图7A)。PG6 在 CD4+和T调节细胞群中富集,而 PG5 细胞在基质细胞群中富集(图 7A)。为了更高分辨率地研究 PG 中的 T 细胞群,基于 43 例病例的scRNA-seq数据,对 T 细胞进行了基于图的聚类分析,得到了 15 个不同的 T 细胞聚类,并根据最近建立的 B 细胞非霍奇金淋巴瘤 T 细胞群汇编对这些细胞簇进行了注释。这些细胞簇包含初始型和(中央/效应)记忆型 CD4+或 CD8 + T 细胞。T细胞、调节性T细胞和滤泡辅助性T细胞,并进一步区分了耗竭和非耗竭的T细胞表型(图7B-C)。在所有PG中,PG6的T细胞绝对计数最高,而PG3和PG4肿瘤的T细胞数量均极少(图7B-C)。大多数T细胞亚群在PG4中显著减少(图7D)。然而,尽管频率较低,某些T细胞亚群在PG4中占主导地位,包括CD8+细胞毒性效应记忆(EM)2和EM3亚群,这两个亚群均表现出明显的耗竭表型(图7E)。为了验证这一发现,他们利用来自验证队列DLBCL肿瘤的组织微阵列,进行了基于免疫荧光的T细胞标志物成像分析。与PG6相比,PG4中CD3+ T细胞的频率显著降低(图7F-H)。PG4肿瘤微环境中相对较少的T细胞呈现耗竭表型,其中PD1+ TIM-3+阳性CD3+ T细胞的比例显著升高(图7G-H),PD1 + LAG3+CD3+ T细胞也呈现类似的趋势。值得注意的是,PG4病例中CD68+ PD-L1+巨噬细胞的比例较高,这可能与观察到的T细胞耗竭有关。
近期研究表明,淋巴瘤TME由特定的细胞微环境(CN)组成,这些CN可通过空间转录组学进行识别。他们确定了82例弥DLBCL病例的PG状态,这些病例已通过靶向空间转录组学进行分析。所鉴定的PG的比例及其分子特征反映了发现队列和验证队列的PG分布。重要的是,在对现有CosMx数据进行重新分析后, CN分布显示出显著差异(图7 I)。PG5中CN7的代表性较高,CN7主要由基质细胞类型组成(图7 I),这与整体和单细胞分析的结果一致。相比之下,PG6中CN3的代表性显著升高,CN3富含T细胞和巨噬细胞,这与其免疫富集表型相符(图7 I)。有趣的是,PG4 中分配到 CN6 的细胞比例很高(图 7I),这些细胞主要由恶性 B 细胞组成,但也包含一部分 T 细胞。与其他 PG 相比,PG4 肿瘤中分配到 CN6 的恶性 B 细胞的 MYC 靶基因特征表达显著更高(图 7J)。此外,PG4 中 CN6 相关的 T 细胞的 T 细胞耗竭特征评分高于非 PG4 肿瘤中的 T 细胞(图 7J)。因此,空间转录组分析验证了 MYC 的细胞内在高活性以及 PG4 的肿瘤微环境特征,表明这些肿瘤具有免疫抑制性TME。
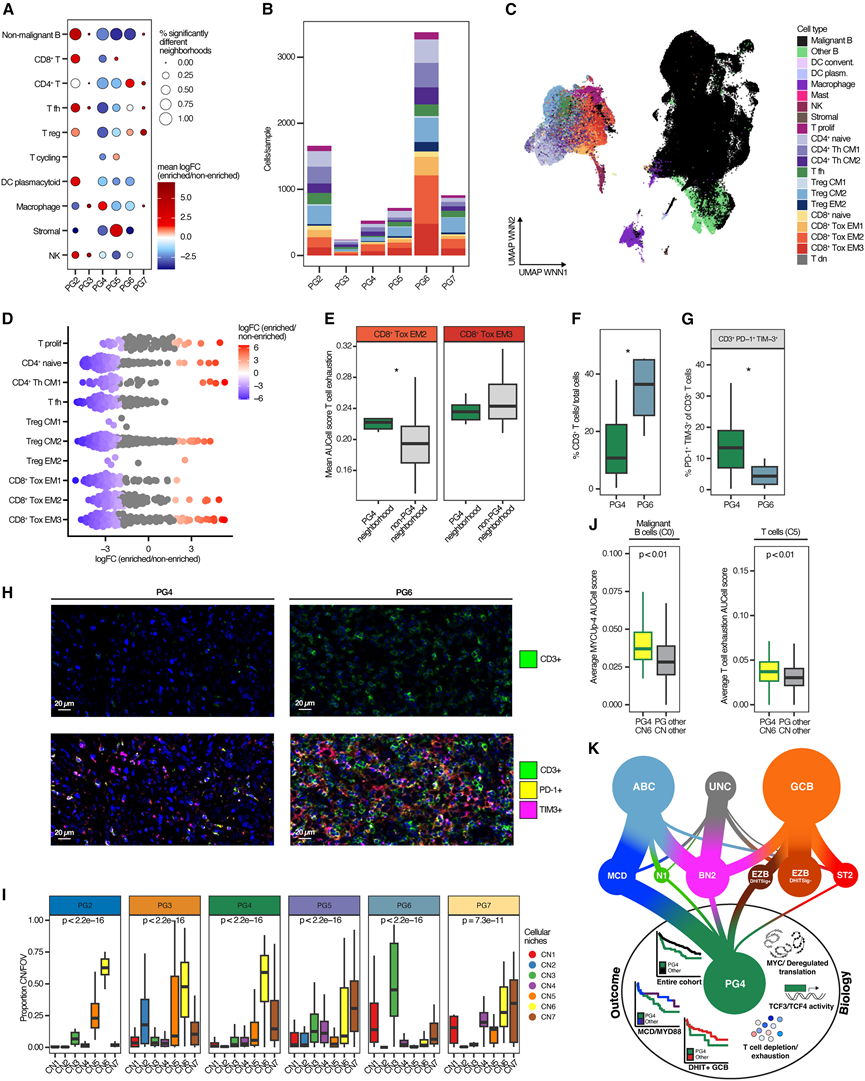
图7. DLBCL蛋白基因型的TME表型。
(A) 不同细胞类型在DLBCL各蛋白基因型中细胞邻域的差异丰度。(B) 按 DLBCL 蛋白基因型分层的每个样本中 T 细胞亚群的绝对数量。(C)测序数据的加权最近邻整合维度的 UMAP 。(D) 将推断出的邻域分配到其主要的 T 细胞类型。(E) 平均AUCell评分。(F) 在验证队列中,通过多重免疫荧光检测DLBCL组织微阵列。(G) 现在计算所有CD3 + T细胞中PD-1+和TIM-3 +细胞的百分比。(H) 代表性多重免疫荧光图像。(I) 根据 CosMx 空间转录组分析确定的 DLBCL 蛋白基因型。(J) PG4 预测病例和部分 CN6 病例中恶性 B 细胞(C0-Tumor-B)每个视野的平均 MYC 靶基因表达评分。(K) PG4-DLBCL 临床和生物学特征的示意图。
+ + + + + + + + + + +
结 论
通过整合来自478例DLBCL肿瘤的多组学数据,本研究鉴定了7种DLBCL的PG,这些基因型反映了涵盖已知分子亚型的特定病理生理特征。PG4与不良预后相关,且这种相关性独立于已知的风险因素,例如细胞起源、国际预后指数或遗传特征。PG4包含活化B细胞样和生发中心B细胞样肿瘤以及基因未分类的病例。它具有暗区相关的B细胞表型,并富集可激活MYC的BTG1突变。多组学分析显示,无论是否存在MYC易位,MYC和TCF3/4的转录活性均增强。PG4肿瘤微环境的特征是CD8+ T细胞耗竭。研究发现了高危 DLBCL 肿瘤的常见致癌主题,并为未来的诊断和治疗方法提供了蛋白质基因组学框架。
+ + + + +



