

 English
English文献解读|Sci Adv(11.7):唐氏综合征的免疫代谢相互作用的综合分析
✦ +
+
论文ID
原名:Integrated analysis of immunometabolic interactions in Down syndrome
译名:唐氏综合征的免疫代谢相互作用的综合分析
期刊:Science Advances
影响因子:11.7
发表时间:2024.12.13
DOI号:10.1126/sciadv.adq3073
背 景
唐氏综合征 (DS) 是人类最常见的染色体疾病,美国大约每 700 名新生儿中就有 1 名患有 21 三体综合征 (T21),而 T21 正是 DS 的诱因。1979 年至 2003 年间,美国每 10000 名活产婴儿中 DS 的发病率从 9.0 增加到 11.8。在同一时期,由于医疗保健的改善,DS 患者的平均寿命从 30 岁增加到 60 岁。尽管寿命有所增加,但 DS 患者及其家人仍面临着并发疾病带来的持续健康挑战,这些疾病在 T21 患者中更为普遍。与一般人群相比,唐氏综合征患者罹患听力和视力障碍、肥胖、血脂异常、先天性心脏缺陷、白血病、多种自身免疫性疾病、阿尔茨海默病以及自闭症和癫痫等神经系统疾病的风险更高。此外,唐氏综合征患者会因病毒性呼吸道感染而出现更严重的并发症,这使他们面临不良预后的风险很高,严重急性呼吸综合征冠状病毒 2 感染就是这种情况。虽然已知与唐氏综合征相关的临床疾病,但人们对 21 号染色体三倍化如何改变分子通路、细胞功能和整体生理学,从而导致观察到的并发疾病模式的机制知之甚少。
实验设计
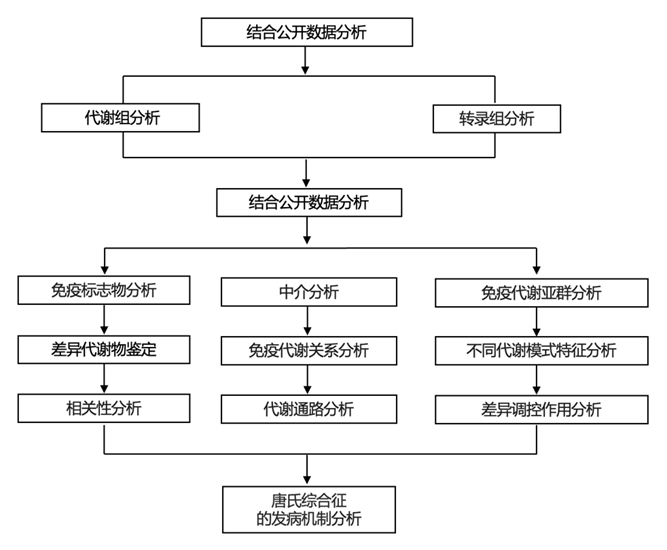
结 果
01

唐氏综合征患者的免疫代谢特征与一般人群不同
为了研究唐氏综合征患者的免疫代谢失调及其与该人群临床特征的关系,研究团队分析了人类三体计划 (HTP) 队列研究生成的数据集。从全血样本生成多组学数据集,包括使用54 种血浆免疫标志物、通过超高效液相色谱-质谱法 (UHPLC-MS) 检测的 174 种血浆代谢物(代谢组学数据),以及通过转录组分析 (RNA-seq) 检测的 12560 个蛋白质编码基因转录本计数。还通过参与者/护理人员调查和医疗记录审查相结合的方式收集了人口统计数据和共病信息。这里呈现的结果来自 290 个个体的子集分析,其中 244 个个体为 21 号染色体的三体(T21),46 个个体为整倍体对照(D21)(图 1A)。
他们对免疫标志物和代谢物进行了差异丰度分析。与以前的报告一致,平均而言,唐氏综合征患者的细胞因子丰度增加,包括促炎细胞因子[例如胸腺基质淋巴细胞生成素 (TSLP)、白细胞介素 (IL)-17D、IL-17C、IL-16 和 IL-6] 和趋化因子[例如单核细胞趋化蛋白-1 (MCP-1) 和干扰素-γ 诱导蛋白 10 (IP-10)],以及急性期蛋白(例如 CRP)。抗炎细胞因子(例如 IL-10 和 IL-1RA)和其他免疫介质(血管内皮生长因子 A、胎盘生长因子和成纤维细胞生长因子)也升高。唐氏综合征患者中显著减少的细胞因子包括肿瘤坏死因子-β (TNF-β) 和 IL-12/IL-23p40。唐氏综合征患者中增加的代谢物包括炎症标志物[例如鞘氨醇、尿酸、5(S) 羟基二十碳四烯酸 (HETE)、12(S)-HETE 和 11-HETE]、色氨酸途径代谢物(例如犬尿氨酸和 5-羟基吲哚乙酸)、胆汁酸(例如牛磺石胆酸和熊去氧胆酸)和溶血磷脂酸 (LPA)(例如 LPA 16:1)。唐氏综合征患者的氨基酸(例如甘氨酸、组氨酸、丝氨酸和酪氨酸)减少。总体而言,代谢物和细胞因子的结果与之前报告的结果一致,并证明数据集是可靠的,并且代表了 DS 患者的生物学特性。
为了确定唐氏综合征患者的免疫代谢失调模式,他们计算了所有细胞因子和代谢物对之间的成对 Spearman 相关性。在患有 T21 的个体中,他们使用k均值聚类(k = 4)确定了具有相似失调模式的细胞因子和代谢物组(图 1B-C)。在细胞因子中,IL-6/CRP/SAA 信号轴中的因子彼此之间显示出显著的正相关性(图 1C)。干扰素-γ (IFN-γ)、IFNL1 (IL-29) 和 IFN 诱导蛋白 IP-10 也呈正相关,并与多种单核细胞活化细胞因子和趋化因子相关,例如巨噬细胞炎症蛋白-1α (MIP-1α)、MCP-1 和 MCP-4。在代谢物中,他们观察到先前观察到在 DS 中失调的色氨酸分解代谢物(如犬尿氨酸和 5-羟基吲哚乙酸)之间存在很强的相关性(图1C)。与 DS 患者密切相关的其他代谢物例子包括白三烯 B4 与十六碳烯酸以及消退素 D1/D2 与保护素 D1。
从免疫标志物和代谢物的综合分析中发现了一些有趣的模式。例如,CRP、SAA 和 IL-6 均与 DS 中的氨基酸水平呈负相关,包括丝氨酸、天冬酰胺和苏氨酸(图 1D)。如前所述,TNF-α 与色氨酸分解代谢物犬尿氨酸和 5-羟基吲哚乙酸呈正相关。他们进一步评估了按代谢类别富集细胞因子和代谢物之间的显著相关性。氨基酸与几种细胞因子呈负相关,包括 CRP、IL-1RA、IL-22、IL-6 和 MIP-1α(图 1E)。脂肪酸/二十烷酸与IL-5和VCAM-1的富集也呈负相关。细胞因子IL-1α与精氨酸和脯氨酸代谢以及吲哚和色氨酸代谢物呈正相关。
为了评估核型之间的差异,他们比较了 T21 患者与整倍体对照 (D21) 患者的细胞因子和代谢物之间的成对相关系数。总体而言,与各组的样本量一致,他们观察到 T21 患者)634 个显著相关)的细胞因子和代谢物之间的相关性比 D21 整倍体对照(91 个显著相关)更显著(图 1F)。为了解决不同核型样本量不平衡的问题,他们随机将 T21 队列再抽样至 46 人,以匹配 D21 队列的样本量。与整个队列一样,与整倍体对照相比,他们在 T21 患者中观察到更显著的细胞因子-代谢物相关性,包括正相关性和负相关性。此外,他们观察到与 T21 队列相比,整倍体对照的相关系数范围较小。例如,在 T21 患者中,他们观察到与对照组相比,细胞因子与犬尿氨酸和N -乙酰神经氨酸之间的正相关性更高(图 1F)。N-乙酰神经氨酸是哺乳动物唾液酸的主要形式,与小鼠的免疫信号传导和识别记忆障碍有关。总之,这些结果揭示了唐氏综合征患者的免疫代谢失调与整倍体个体不同且更具异质性,这可能是该人群中关键并发疾病的病因之一。
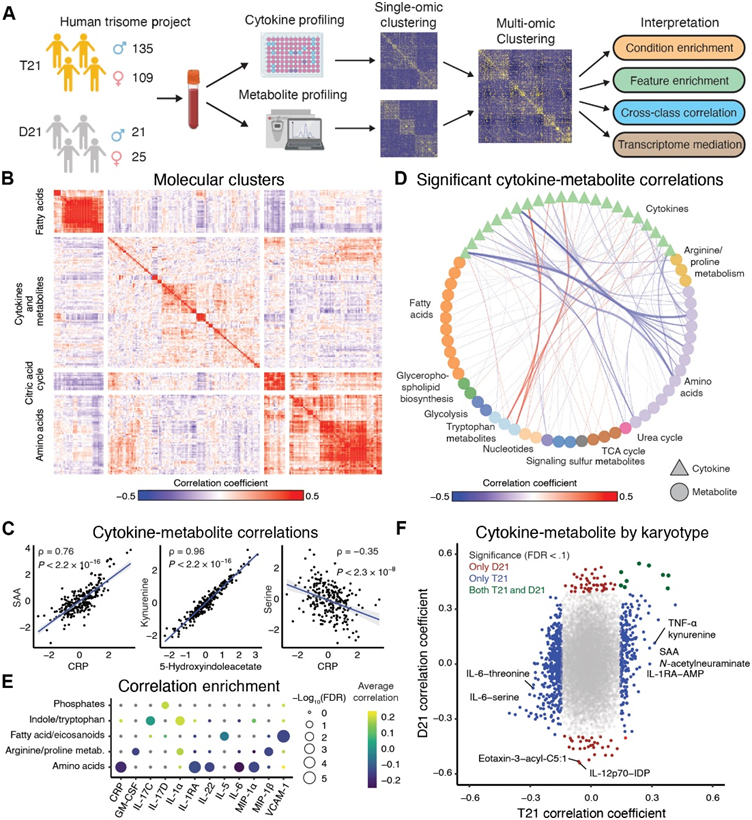
图1. DS的免疫代谢特征与一般人群不同。
(A)实验流程。(B) 标准化细胞因子和代谢物丰度之间的 Spearman 相关系数热图。(C) 细胞因子、代谢物之间以及跨分子检测类型标准化值的代表性散点图。(D) 按代谢物类别汇总的细胞因子和代谢物之间的统计学显著 (FDR < 0.1) 相关性。(E) 代谢物类别的细胞因子和代谢物之间显著相关性。(F) T21 和 D21 人群中细胞因子和代谢物之间的 Spearman 相关系数。
02
中介分析揭示了驱动 DS 中细胞因子-代谢物关系的信号通路
细胞因子可通过改变参与代谢级联的酶的调节和表达来间接影响代谢物水平。例如,IFN-γ信号通过IFN-γ受体IFNGR1/IFNGR2激活Janus激酶1和2(JAK1/2),后者又磷酸化并激活信号转导和转录激活因子1(STAT1)转录因子(图2A)。STAT1 转位至细胞核后,诱导数百个 IFN 刺激基因 (ISG) 的转录,包括IDO1 ,它编码吲哚胺 2,3-双加氧酶,这是色氨酸分解代谢途径中色氨酸转化为犬尿氨酸的限速酶。I型和 III 型 IFN 也通过类似途径诱导IDO1表达。
为了预测这些类型的中介关系,他们使用 IFN-γ(II 型 IFN)作为阳性对照,使用匹配的转录组数据来识别可以调节细胞因子-代谢物关系的基因。使用偏相关性,证明STAT1介导 IFN-γ 和犬尿氨酸之间的关系,这是因为当关系以STAT1为条件时,相关性下降了 88% (图 2B)。该结果表明,通过偏相关中介分析来分析细胞因子、代谢物和基因表达测量值可以捕捉已知的生物调控,并可用于优先考虑基因作为免疫代谢关系的潜在介质。
为了评估潜在基因调控的特异性,他们对所有其他免疫代谢关系进行了中介分析(图 2C)。他们确定了几种已知基因作为 IFN-γ-犬尿氨酸关系的调节剂。总体而言,STAT1是排名第六的基因(图 2D)。鸟苷酸结合蛋白 2 和 1(GBP2/1)分别排名第一和第八。敲低 GBP 可抑制人间充质基质细胞中IDO1 的表达。另一个排名靠前的潜在介质是 Fcγ 受体 I(FCGR1A)。IFN-γ 可增加FCGR1A 的表达和 FCGR1A 对免疫复合物的相对亲和力。相反,与已知关系一致,STAT1最显著介导的免疫代谢关系是 IFN-γ 与饱和脂肪酸(例如十二烷酸、二十二碳五烯酸和十四烷酸)或色氨酸分解代谢物(例如犬尿氨酸和 5-羟基吲哚乙酸)之间的关系(图 2E)。
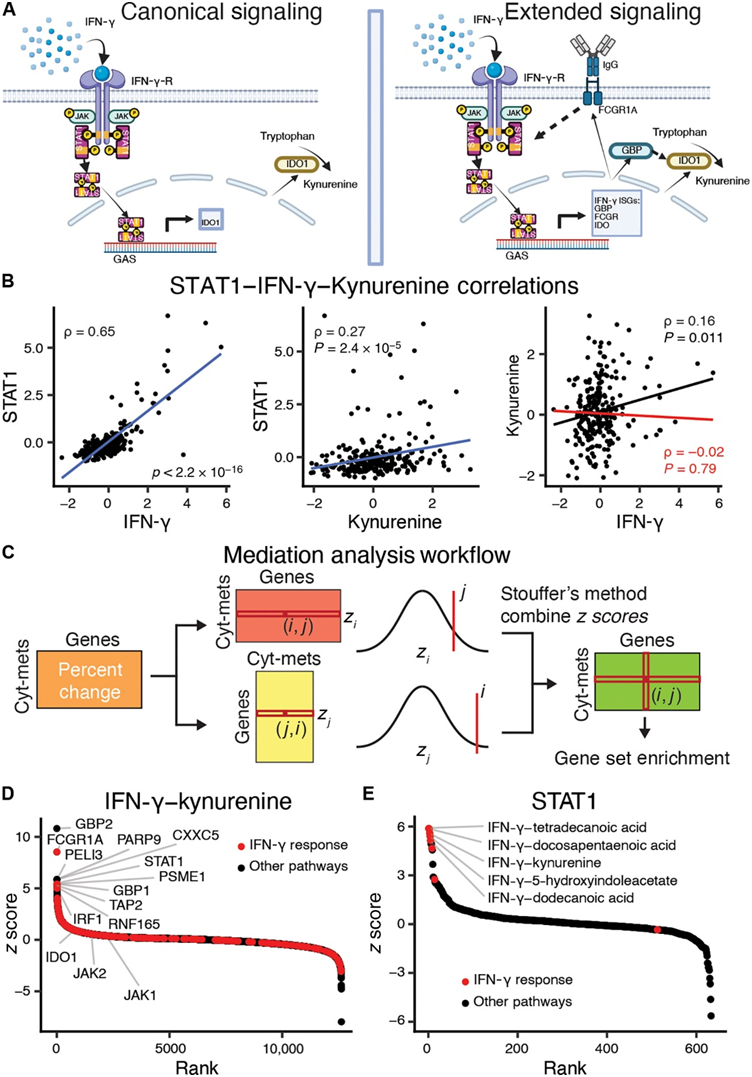
图2. 细胞因子诱导基因表达变化,从而影响代谢物水平。
(A)机制示意图。(B) IFN-γ–STAT1、犬尿氨酸-STAT1 和 IFN-γ-犬尿氨酸的散点图。(C) 识别以基因表达为条件的免疫代谢关系。(D) IFN-γ-犬尿氨酸的基因中介排名,其中 IFN-γ 反应基因以红色突出显示。(E) STAT1介导的细胞因子代谢物排名,其中富集 IFN-γ 反应基因的细胞因子代谢物关系以红色突出显示。
03
不同的转录程序介导免疫代谢关系
为了了解转录介导对细胞途径和过程的影响,他们对每个细胞因子-代谢物关系的介导基因z分数的排序列表进行了基因集富集分析。之前已确定全血转录组中基因表达的 T21 特异性变化,包括 IFN-γ 反应、血红素代谢途径和氧化磷酸化的激活。根据细胞因子-代谢物关系中标准化富集得分 (NES) 的方差对基因集进行排序,并通过途径介导 NES 对细胞因子-代谢物关系进行层次聚类,结果显示,IFN-γ 和 IFN-α 反应途径在细胞因子-代谢物关系中差异很大。氧化磷酸化和血红素代谢途径介导的细胞因子-代谢物关系与 IFN-γ 和 IFN-α 关系截然不同。总之,这些结果表明,在唐氏综合征患者中,特定的细胞因子-代谢物关系是由截然不同的转录程序介导的。
IFN-γ 反应基因在 93 种细胞因子-代谢物关系中显著富集 (FDR < 0.05) (图 3A)。在根据 IFN-γ 反应富集分数对细胞因子-代谢物关系进行排序时,他们观察到最高的结果是 IFN-γ 和十二烷酸(或月桂酸)(图 3B)。月桂酸已证明能抑制 IFN-γ 对巨噬细胞中细胞间粘附分子 1 (ICAM-1) 和血管细胞粘附分子 1 (VCAM-1) 表达的影响。IFN-γ 和十二烷酸之间的相关系数确实为负相关,表明这种关系可能受 IFN-γ 反应基因调节。总体而言,IFN-γ 的代谢通路富集分析显示,在免疫代谢相互作用中,糖(如麦芽糖、甘露醇、d -核糖、d -鼠李糖和d -阿拉伯糖醇)和饱和脂肪酸(例如己酸、庚酸、辛酸、壬酸、癸酸、十二酸和十四酸)显著富集。
血红素代谢基因在介导89种细胞因子-代谢物关系中显著富集(图3A)。多巴胺与细胞因子 MCP-4 和嗜酸细胞活化趋化因子之间的免疫代谢关系最富集于血红素代谢介导(图 3D)。在细胞因子 ICAM-1、巨噬细胞衍生细胞因子 (MDC)、IFN-β 和 VCAM-1 与胆汁酸(例如熊去氧胆酸和鹅去氧胆酸)之间观察到了其他排名靠前的细胞因子-代谢物关系。对由血红素代谢介导而富集的免疫代谢关系进行代谢物途径富集分析表明,最具代表性的代谢物类别包括氨基酸和胆汁酸。
氧化磷酸化基因在介导80种细胞因子-代谢物关系方面显著富集(图3A)。炎症细胞因子IL-6与尿素循环代谢物鸟氨酸之间的关系最显著地富集于氧化磷酸化介导(图3C)。此外,他们观察到氧化磷酸化介导了几种涉及精氨酸和脯氨酸代谢产物(例如鸟氨酸、脯氨酸、反式-4-羟基-L-脯氨酸、4-5-胍基-2-氧代戊酸)的免疫代谢关系。他们观察到免疫代谢关系主要受 DS 中失调的不同转录程序调控,例如 IFN-γ 反应、血红素代谢和氧化磷酸化(图 3E)这些结果表明,DS 中失调的通路会影响独特的免疫代谢关系。
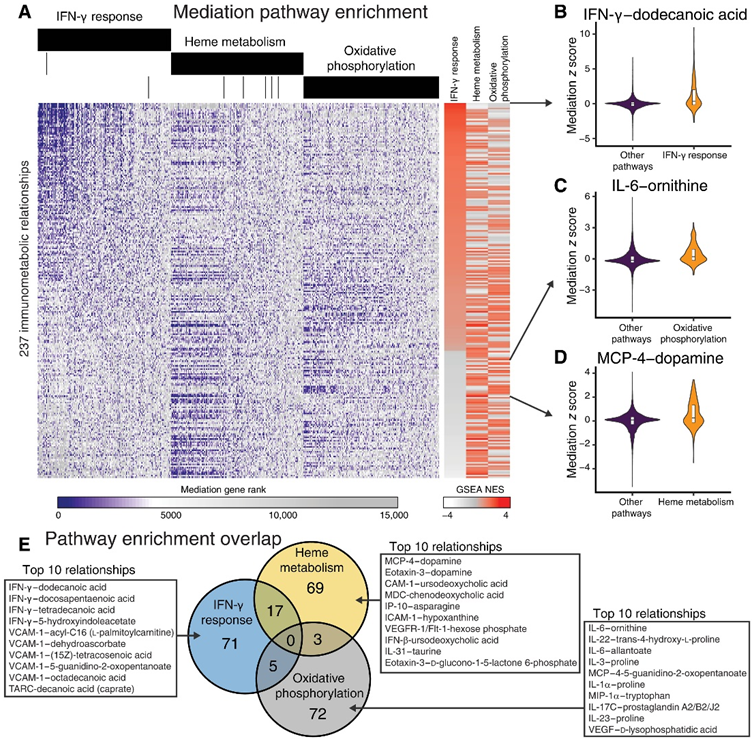
图3. 中介分析揭示了 DS 中驱动细胞因子-代谢物关系的信号通路。
(A) 所有基因在细胞因子-代谢物关系中的基因介导排名。(B-D) 最富集 (B) IFN-γ 反应、(C) 氧化磷酸化或 (D) 血红素代谢介导的免疫代谢关系的介导分数。(E) 细胞因子-代谢物关系的重叠。
04
整合免疫标志物和代谢物可识别唐氏综合征的临床亚组
他们确定了四个亚组的综合免疫代谢聚类,这些亚组具有不同的免疫代谢相互作用和临床特征(图 4A- B)。他们将综合亚组与基于个体细胞因子或代谢物谱的亚组进行了比较,后者分别由两个和三个亚组组成。综合解决方案将队列分层为独特的组,结合了来自单组学聚类解决方案的个体(图 4C),这表明整合数据可以识别单组学谱中未捕获的分子模式。此外,与单组学数据相比,基于整合多组学数据的聚类之间存在更丰富的共生条件(图 4A-C),他们将整合的聚类称为免疫代谢亚群 (IMS) 1-4。
免疫代谢亚群 1 (IMS1) 由 71 名个体组成。与其他 T21 患者相比,该组患者中患有自身免疫性皮肤病史的人较多(图 4B)。IMS1 中的个体具有更高的脂肪酸/二十烷酸水平。例如,十六烷酸(棕榈酸)是西方饮食中普遍存在的饱和脂肪酸,该亚组中的十六烷酸(棕榈酸)含量较高(图 4D)。十六烷酸与适应性免疫有关,并且已知它通过诱导神经酰胺代谢来增强 Toll 样受体依赖性炎症。
IMS2 由 91 名个体组成,该亚群中肥胖个体和有频繁/复发性肺炎病史的个体代表性不足(图 4B)。该亚组中氨基酸含量较高,而脂肪酸/二十烷酸含量较低。IMS2 中升高最显著的特征是乙酰胆碱(图 4E)。乙酰胆碱通过免疫细胞上的α7烟碱乙酰胆碱受体抑制炎症细胞因子的释放和炎症,这表明乙酰胆碱可能减轻该亚组中的炎症作用。
IMS3 由 31 名个体组成,该亚群富含有甲状腺功能减退症、抑郁症和肥胖病史的个体(图 4B)。该亚组中有自身免疫性皮肤病史的人也较少。该亚组中的个体往往具有更高的炎症细胞因子丰度(例如 IFN-γ、SAA 和 IL-29)(图 4F)和色氨酸分解代谢物(如犬尿氨酸和 5-羟基吲哚乙酸)(图 4G)该聚类还富含较低丰度的脂肪酸/二十烷酸。
IMS4 由 43 名个体组成。该组人群中,有频繁或复发性肺炎病史的人群较多,有自身免疫性皮肤病病史的人群较少(图 4B)。该组中的个体氨基酸水平明显较低。例如,与所有其他具有 T21 或 D21 核型的个体相比,IMS4 中的丙氨酸含量明显较低(图 4H)。
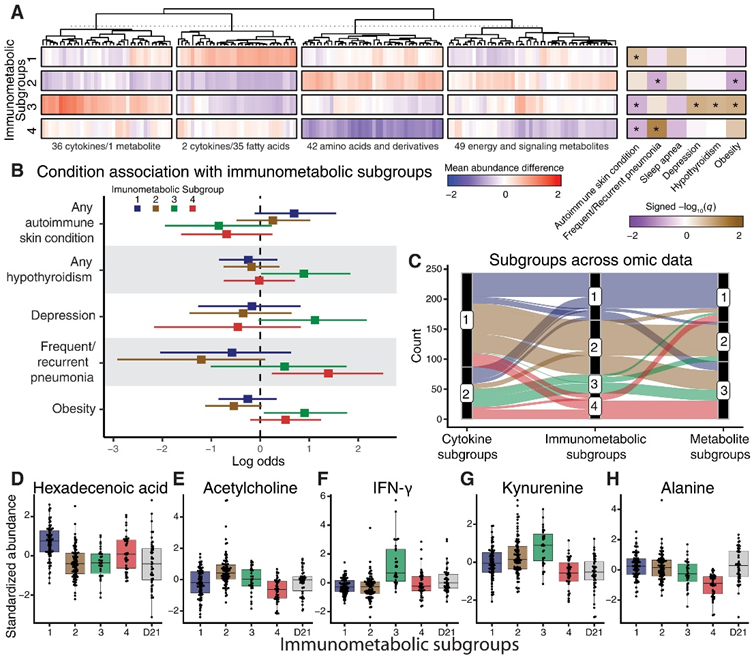
图4. 免疫代谢关系定义了 DS 的临床亚组。
(A) 按免疫代谢亚组划分的平均特征丰度(IMS 与所有其他)的层次聚类差异。(B) 与每个免疫代谢亚组相关的报告共现条件的对数优势比。(C) 冲积图显示基于细胞因子谱、代谢物谱或免疫代谢谱的个体在亚组中的分布。(D-H) 免疫代谢亚组中单个细胞因子或代谢物的标准化丰度以及与具有 D21 核型的个体的关系。
05
免疫代谢亚群揭示了细胞因子-代谢物关系的差异介导
相比之下,IMS1(自身免疫性皮肤病病史增多)和IMS3(甲状腺功能减退、抑郁和肥胖病史增多)表现出DNA修复基因组介导的免疫代谢增强(图5A)。相反,IMS4(频繁/复发性肺炎病史增加)显示参与血管生成的基因介导增加。IMS2 包括具有更健康临床特征的人,其 Hedgehog 信号富集和免疫代谢介导相对较高。在四个 IMS 中,IFN-γ、血红素代谢和氧化磷酸化基因组富集免疫代谢介导的速率相似。
接下来,他们研究了 IMS 中差异最大的三种通路,具体为 DNA 修复、Hedgehog 信号传导和血管生成(图 5A)。他们比较了通路介导富集的代谢物类别,发现即使免疫代谢特征的总体数量同样富集,也存在共同和不同的免疫代谢介导类别。例如,在 IMS1 和 IMS3 中,涉及氨基酸的免疫代谢关系都因 DNA 修复基因的介导而富集;然而,与脂肪酸/二十烷酸的关系在 IMS1 中明显由其介导,而与吲哚和色氨酸代谢物的关系在 IMS3 中明显由其介导(图 5B)。
为了进一步研究这一点,他们确定了这些代谢物聚类中的免疫代谢关系和最重要的基因介导关系。对于 IMS1 中的 DNA 修复基因,POLR2J(RNA 聚合酶 II 亚基 J)是 VCAM-1 和二十烷酸(例如前列腺素 A3/B3 和白三烯 B4/PGA1/PGB1)之间几种关系的主要介导者(图 5C)。IMS2 揭示了 Hedgehog 信号通路介导的涉及氨基酸的免疫代谢关系,其中极低密度脂蛋白受体(VLDLR)是涉及氨基酸(例如甘氨酸)的几种免疫代谢关系的主要介导者(图 5D)。在IMS3中,肌苷酸脱氢酶2(IMPDH2)是炎症细胞因子(例如CRP、SAA和IL-17C)与色氨酸分解代谢物(犬尿氨酸和5-羟基吲哚乙酸)之间关系的主要介质(图5E)。在有频繁/复发性肺炎病史的人群中富集的IMS4中,一些免疫代谢关系是由血小板衍生的生长因子亚基A (PDGFA)介导的(图5F)。

图5. 免疫代谢亚群揭示了细胞因子-代谢物关系的差异介导。
(A) 在每个免疫代谢亚群 (IMS) 的基因集中,基因介导的免疫代谢关系显著 (FDR < 0.05) 和正向富集 (NES > 0) 的数量。(B) IMS 之间差异最大的基因集介导的细胞因子-代谢物关系的代谢物类别的富集。(C-F) 报告了每个 IMS 中选定代谢物类别中细胞因子-代谢物关系的基因介质及其相对强度。
+ + + + + + + + + + +
结 论
本研究多组学数据,推断出由特定转录程序介导的细胞因子-代谢物关系。与整倍体对照相比,唐氏综合征患者的干扰素反应、血红素代谢和氧化磷酸化基因介导的免疫代谢相互作用有所增加。对唐氏综合征患者免疫代谢关系的无监督聚类揭示了同时出现多种疾病的亚组。在各个亚组中,观察到 DNA 修复、Hedgehog 信号和血管生成的不同介导。分子分层与 DS 中观察到的临床异质性相关,这表明整合多个组学谱可以揭示特定于 DS 共病状况的协调失调轴。
+ + + + +



