

 English
English文献解读Cell Rep Med(10.6):儿童高级别胶质瘤的纵向单细胞和空间多组学图谱
✦ +
+
论文ID
原名:A longitudinal single-cell and spatial multiomic atlas of pediatric high-grade glioma
译名:儿童高级别胶质瘤的纵向单细胞和空间多组学图谱
期刊:Cell Reports Medicine
影响因子:10.6
发表时间:2026.04.23
DOI号:10.1016/j.xcrm.2026.102766.
背 景
儿童高级别胶质瘤(pHGG)是一种极其严重的脑部恶性肿瘤,约占婴幼儿至青少年中枢神经系统(CNS)肿瘤的11%。虽然pHGG的发病率相对较低(每10万人口1.78例),但预后极差,中位总生存期仅为10-18个月。虽然经过数十年的研究和超过1500项胶质瘤临床试验,但目前仍无法治愈pHGG。标准治疗方案包括最大限度的安全切除、大剂量放疗和化疗,但这种多模式治疗仅能将总生存期延长数月。虽然儿童和成人高级别胶质瘤(HGG),包括多形性胶质母细胞瘤(GBM),在组织病理学和临床特征方面有很多共同之处,但基因组学、转录组学和表观基因组学数据存在差异,单细胞多组学和空间分析技术的最新进展极大地增进了大众对成人和儿童脑肿瘤肿瘤内和肿瘤间异质性的理解。
实验设计
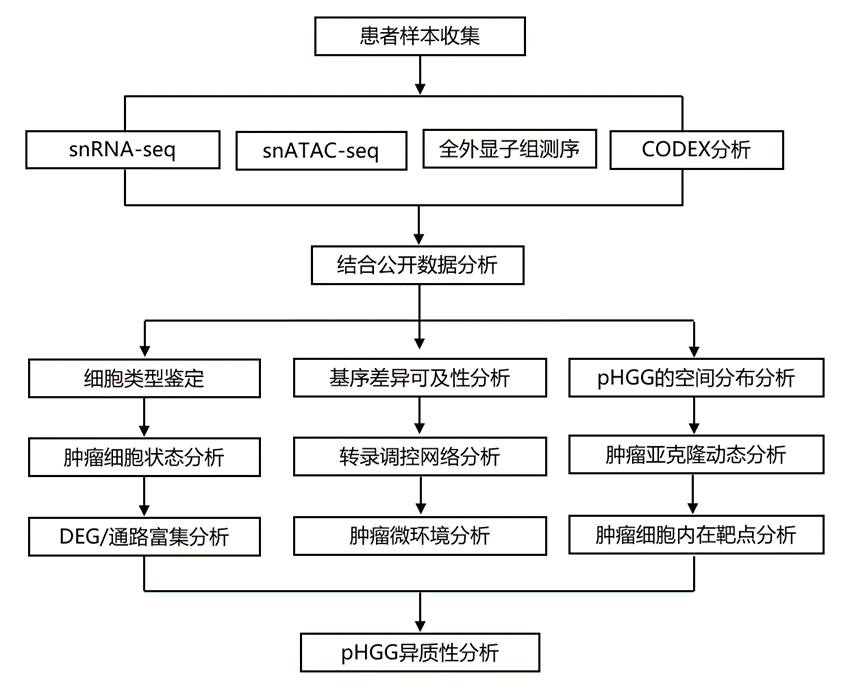
结 果
01

纵向pHGG样本的单细胞分析
研究团队对来自儿童脑肿瘤网络 (CBTN)的患者的 pHGG 样本进行了分析,这些样本涵盖了不同的治疗时间点。分析方法包括单细胞核转录组分析(snRNA-seq)(15 对样本)和单细胞核转座酶可及染色质测序 (snATAC-seq)(11 对样本)(图1A-C)。纳入标准为:经组织学诊断为高级别胶质瘤且年龄小于 18 岁的患者,将这些患者前瞻性地纳入研究,以便在复发、进展或尸检时进行后续分析。所有患者均接受了放射治疗和手术切除,部分患者还接受了药物治疗,包括替莫唑胺、免疫疗法(例如帕博利珠单抗)和细胞毒性化疗。该队列患者确诊时年龄为4至18岁,男女比例为2.2,肿瘤表现出多种基因组改变。肿瘤标本取自多个解剖部位,包括皮质叶和中线结构(丘脑和小脑)。该队列包括3例H3K27M突变病例、2例H3G34R/V突变病例和1例儿童IDH1突变病例;其余病例均为IDH1/H3野生型(WT)(图1D)。甲基化谱分析将12例患者归类为“儿童型弥漫性高级别胶质瘤”,2例患者归类为“成人型弥漫性胶质瘤”,2例患者归类为其他类别。总共通过snRNA-seq分析了超过40万个细胞,并在质量评估和过滤后通过snATAC-seq分析了超过11万个细胞,分别平均每个细胞捕获了2280个基因和19094个独特的染色质片段。他们捕获了胶质瘤中存在的主要细胞类型,包括正常成熟神经元和少突胶质细胞、髓系细胞(巨噬细胞/小胶质细胞)、T细胞、内皮细胞、壁细胞,以及他们称之为“其他神经元和胶质细胞”的多种细胞群,其中包括推测的肿瘤性和非肿瘤性亚群的混合物(图1B-C)。患者和时间点之间存在显著的异质性(图1D)。值得注意的是,大部分壁细胞来自两名患者,而T细胞主要来自一名患者(图 1D)。对snRNA-seq 数据中细胞类型组成的纵向变化进行分析发现,在患者匹配对中,非肿瘤性少突胶质细胞和成熟神经元显著增加(图1E)。snATAC-seq 数据也显示,治疗后少突胶质细胞显著富集(图1F),这与先前在成人胶质母细胞瘤 (GBM) 中的观察结果一致。
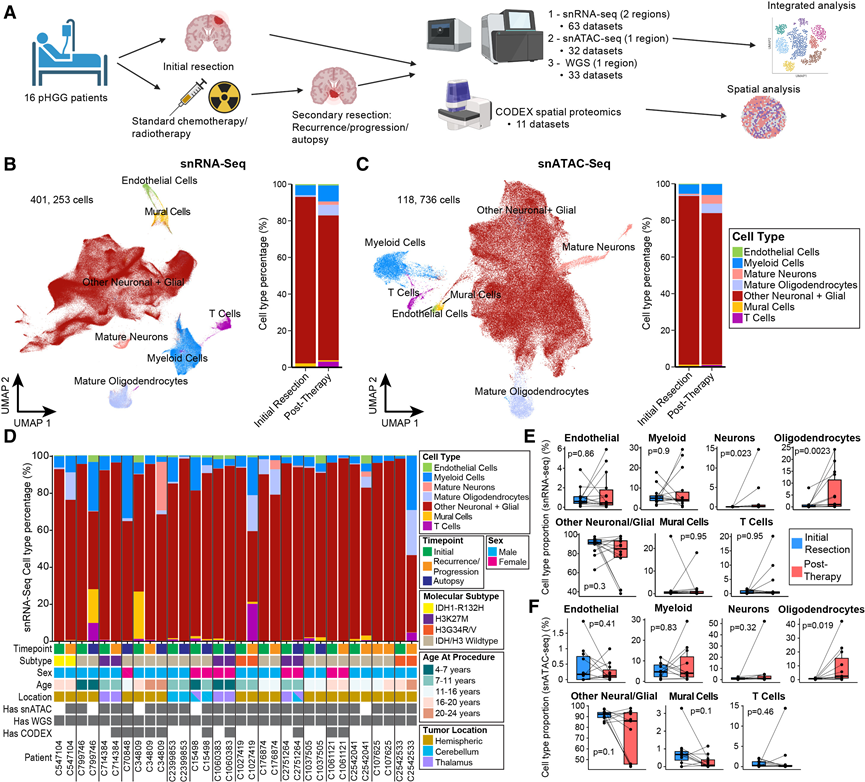
图1. 儿童高级别胶质瘤 (pHGG) 的纵向单细胞 RNA 和 ATAC 图谱。
(A) 患者匹配的纵向pHGG标本的多组学研究概述。(B-C) 均匀流形近似和投影(UMAP),按主要细胞类型类别进行注释,以及堆叠条形图,比较初始切除的 pHGG 样本与治疗后样本的整个数据集的细胞类型比。(D) 每个患者和治疗时间点的 snRNA-seq 数据中的细胞类型比例。(E-F) 分别展示了每位患者在初次切除和治疗后时间点细胞类型比例的变化。
02
儿童神经胶质瘤表现出不同的肿瘤细胞状态
他们旨在表征肿瘤细胞群,并评估这些细胞状态在疾病进展和治疗过程中的变化。为此,通过拷贝数变异(CNV)推断识别出疑似肿瘤细胞,并发现治疗后恶性细胞的比例显著降低。随后,他们重新整合了这些细胞群,并通过对整合数据进行无监督聚类,识别出一组核心的pHGG恶性细胞状态。他们基于这些细胞状态的差异表达基因和通路、已发表的源自H3K27M突变和IDH/H3野生型成人和儿童HGG的胶质瘤细胞状态特征的相对表达,以及在发育中胎儿脑参考图谱上的投影,对这些细胞状态进行了注释(图2A-G)。首先,他们鉴定出一种间充质样(MES)状态,该状态表达已知的间充质标志基因(例如CD44、VIM、ANXA1和NDRG1)和血管生成基因(例如VEGFA),并且富含缺氧反应特征(图 2D-E)。此外,他们观察到,虽然间充质细胞的缺氧程度最高,但不同样本之间,甚至在同一细胞状态下,缺氧程度也存在很大差异(图2H)。有趣的是,虽然间充质细胞状态是近期才在 H3K27M 突变型胶质瘤中发现的,但他们在唯一一例儿童 IDH 突变型胶质瘤病例中观察到了低频的 MES 样状态(图 2C)。接下来,他们鉴定出一个细胞群,该细胞群高表达先前定义的少突胶质细胞祖细胞 (OPC) 样和神经祖细胞 (NPC) 样基因特征,并富集于神经发育和突触相关通路,他们将其注释为 OPC/NPC 样细胞群(图 2E)。然后,他们鉴定出两个更分化的细胞群:一个少突胶质细胞 (OC) 样状态,该状态表达成熟少突胶质细胞基因(例如BCAS1和SIRT2),并富集于先前在 H3K27M 突变型胶质瘤中定义的 OC 样特征 (图 2D);以及一个神经元 (NEU) 样状态,该状态高表达一些 NPC 样标记基因(例如SOX4和CD24)。该神经元富含神经递质释放通路,当投射到发育中的人类胎儿脑图谱上时,其形态与兴奋性神经元最为相似(图2G)。NEU 样状态主要局限于几种 IDH/H3-WT 肿瘤(图2C)。
随后,他们在一组IDH-WT成人(n =20)和儿童(n=8)胶质母细胞瘤(GBM)患者中,鉴定出两种高表达星形胶质细胞标志物GFAP的细胞群,它们在Neftel等人建立的星形胶质细胞(AC)样基因模块中富集程度最高(图2D-F)。第一种状态部分投射到神经胶质祖细胞上,而第二种状态最接近成熟星形胶质细胞;因此,他们将这两种状态分别命名为GPC样和AC样。GPC样状态也相对富集Neftel OPC样基因模块,而非Neftel NPC样基因模块,这与GPC的双潜能特性相符(图2G)。为了研究这些细胞状态的分化轨迹,他们对肿瘤细胞进行了RNA速度和伪时间分析。这些分析揭示了以GPC样状态为中心的保守层级动态,其向量场指向星形胶质细胞、少突胶质细胞和神经元转录程序,类似于成人GBM中描述的模式。尽管终末状态的表征在不同患者和亚型之间存在差异,但这些轨迹的总体拓扑结构是可重复的(图2I)。此外,基因和通路分析表明,在星形胶质细胞/间充质状态和前神经元状态之间存在两个过渡状态,并具有中间或混合表型。最后,他们发现了一小群独特的肿瘤细胞,主要存在于单个患者中,这些细胞富含热休克蛋白基因,并注释为HSP+细胞,可能代表尸检诱导的表型;此外,他们还发现了一群在不同患者中共有的增殖细胞(图2C-E)。综上所述,这建立了一个强大的单核转录组肿瘤细胞状态图谱,可用于儿科和成人 HGG 单核数据的参考映射。
这些细胞状态在不同样本中的比例高度异质,患者间和时间点间均存在显著差异(图2C)。然而,对每位患者的肿瘤细胞状态比例进行分析发现,在治疗的不同时间点,包括IDH/H3-WT患者的亚组分析中,细胞类型组成均未发生显著变化(图2J)。相反,他们观察到了异质性的患者特异性轨迹,类似于近期在成人胶质母细胞瘤(GBM)中描述的情况(图2K)。为了验证他们的发现,他们利用snRNA-seq参考数据对包含23对配对样本的201例pHGG患者的更大样本量的RNA-seq数据进行了反卷积分析。首先,基于他们核心的pHGG细胞群,他们发现不同亚型之间的肿瘤细胞状态存在显著差异:GPC样和OC样细胞在H3K27M胶质瘤中最为富集,增殖细胞在H3G34R/V胶质瘤中最常见,而NEU样细胞则主要存在于多个IDH/H3-WT样本中。随后,对配对样本的分析证实了他们之前的发现,即治疗后非肿瘤性少突胶质细胞富集,但未观察到肿瘤细胞群内任何显著的细胞状态转变。
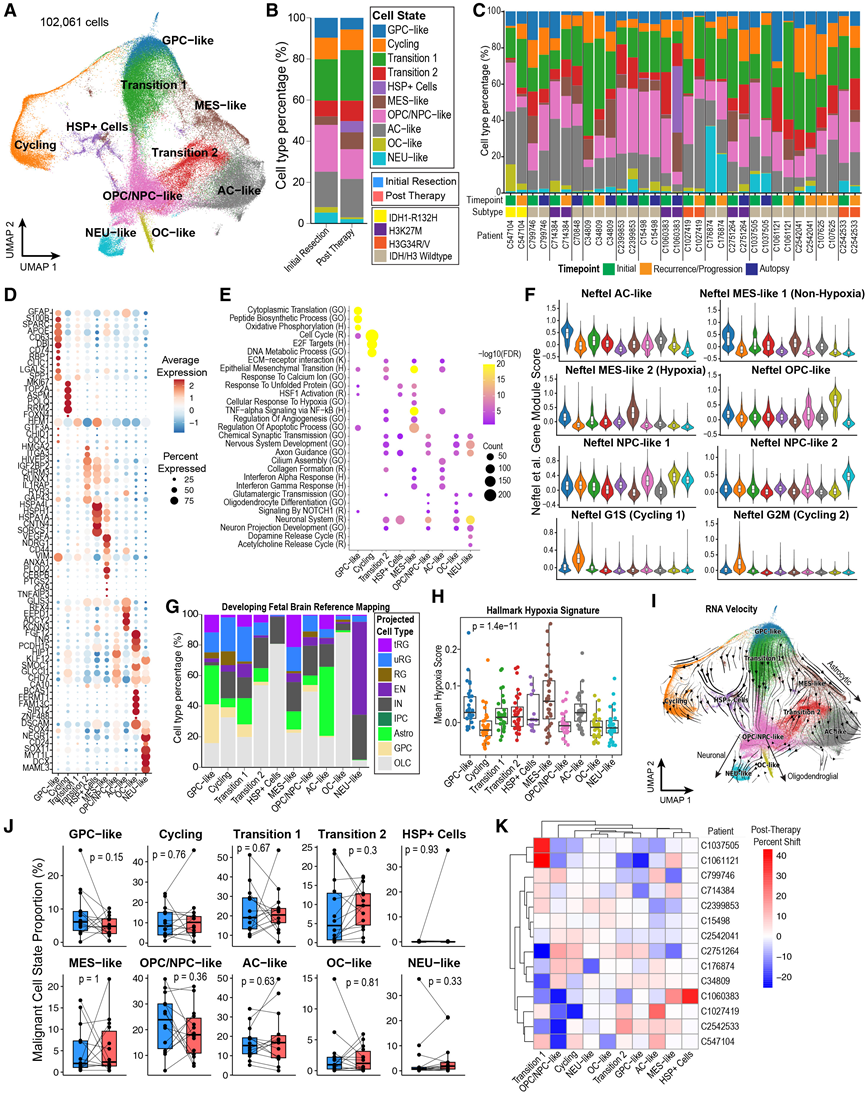
图2. pHGG肿瘤细胞的转录状态。
(A) 整合和注释细胞状态。(B) 比较初始切除和治疗后样本中肿瘤细胞的细胞类型比例的条形图。(C) 比较初始切除和治疗后样本中肿瘤细胞的细胞类型比例的条形图。(D) snRNA-seq 数据中肿瘤细胞状态下代表性差异表达基因的表达。(E) snRNA-seq 数据中肿瘤细胞状态下代表性差异表达基因的表达。(F) GBM细胞状态基因特征的小提琴图。(G) 将每种细胞状态下的细胞类型比例投影到发育中的胎儿人脑数据集。(H) 缺氧特征评分计算为每个细胞状态和每个样本的平均值。(I) 肿瘤细胞的 RNA 速度梯度显示出层级轨迹。(J) snRNA-seq 中每位患者在初次切除和治疗后时间点肿瘤细胞状态比例的变化。(K) 针对每位肿瘤细胞状态和患者,绘制细胞状态转变的热图,颜色代表治疗后样本与初始切除样本之间百分比的差异。
03
转录因子共同调控pHGG肿瘤细胞状态
随后,他们旨在通过单核染色质可及性数据进一步表征这些细胞状态。在将snATAC-seq数据中假定的肿瘤细胞群重新整合并注释后,他们捕获了snRNA-seq数据中转录组定义的所有细胞状态,证实了差异表达基因(DEG)中染色质可及性的富集,并观察到与snRNA-seq在不同样本间具有显著一致性(图3A-B)。正如预期,他们观察到患者和治疗时间点之间存在显著的异质性,且治疗后肿瘤细胞状态未发生显著变化(图3C)。
接下来,他们旨在鉴定调控各细胞状态的转录因子,首先使用chromVAR评估转录因子基序的差异可及性。与之前的报道一致,AP1家族转录因子(例如FOS、JUN和JDP2)的基序在间充质状态下富集(图3D)。值得注意的是,这些基序在过渡2状态下也富集。AC样和GPC样状态均富集RFX因子基序,而OPC/NPC样、OC样和NEU样状态则富集促神经转录因子(例如ASCL1/2、NHLH1和LHX4)(图3D)。
接下来,他们整合了snRNA-seq和snATAC-seq数据,构建了每种细胞状态的转录调控网络(TRN),以预测状态特异性的增强子-启动子相互作用和转录因子-靶基因对。这些TRN揭示了转录因子在调控细胞状态特异性基因表达方面存在显著的协同作用(图3E-G)。RFX因子可调控AC样和GPC样细胞状态。SOX4可通过与其他转录因子(包括LHX1和KLF12)的协同作用,分别调控NEU样和OPC/NPC样状态(图3E-F)。AP1因子和RUNX1预测可共同调控MES样状态。预测RUNX1靶向MES样状态下差异表达最显著的基因,且RUNX1自身在治疗后MES样肿瘤细胞中的表达上调(图3F)。虽然RUNX1已是间质型胶质母细胞瘤(GBM)的促成因子,但这些结果表明RUNX1转录因子是MES样转录状态的核心调控因子。此外,当分别构建IDH/H3-WT和H3K27M亚群的TRN时,RUNX1仍然是核心调控因子。总而言之,该分析揭示了肿瘤表型谱系中重叠的转录调控相互作用。
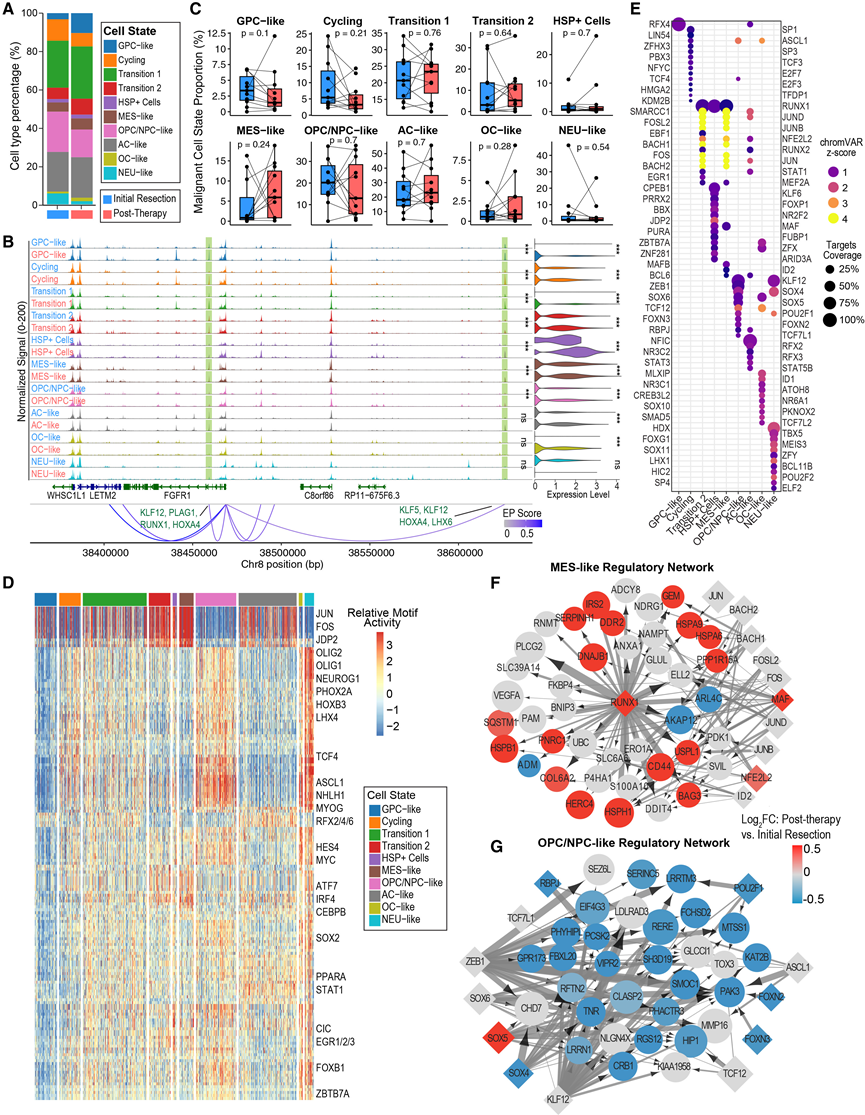
图3. pHGG肿瘤细胞状态的转录调控。
(A) 堆叠条形图,比较初始切除和治疗后样本中肿瘤细胞状态的细胞类型比例。(B) 代表性的ATAC信号轨迹,展示了FGFR1基因座及其在不同细胞状态和时间点的基因表达情况。(C) snATAC-seq 中每位患者在初次切除和治疗后时间点肿瘤细胞状态比例的变化。(D) 各高级别胶质瘤(pHGG)肿瘤细胞状态下差异转录因子(TF)基序可及性的热图。(E) 基于预测的增强子-启动子相互作用和转录因子-靶基因对,概述每种肿瘤细胞状态下最重要的15个转录调控因子。(F-G) 转录调控网络(TRN),分别对应MES样状态和OPC/NPC样状态。
04
肿瘤免疫微环境以多种髓系细胞群为主
在确定了pHGG中的肿瘤细胞状态后,他们着手表征其免疫微环境。T细胞约占snRNA-seq捕获细胞的2%,主要存在于两例治疗后标本中(图1B-1D)。一例进展性H3G34R/V突变病例是一个显著的异常值,其T细胞约占捕获细胞的22%(图1D)。T细胞丰度较低,但异常值高达总细胞组成的20%左右,这与成人GBM的特征相符。髓系细胞约占snRNA-seq数据的9%,且在每位患者中均有捕获,因此他们选择该细胞群进行进一步分析。重新整合后,他们鉴定了11个不同的髓系细胞群,并根据其差异表达基因和转录调控子对其进行了手动注释(图4A-C)。大多数样本包含髓系亚群的分布,而少数样本则以单一亚型为主。此外,这些细胞形成了一个连续的表型谱,包括驻留小胶质细胞和骨髓来源的巨噬细胞的发育过程。
髓系亚群包括组织驻留小胶质细胞、树突状细胞和多种肿瘤相关巨噬细胞 (TAM) 亚群,这些亚群此前已在多种实体瘤类型中得到表征,包括神经胶质瘤。这包括差异表达VEGFA和糖酵解酶(即HK2和ENO2)的促血管生成TAM 、脂质相关 TAM(PPARG和LPL)、炎症性 TAM(NFKB1和IL1B)、干扰素(IFN) 反应性 TAM(IFIT2、IFIT3和ISG15),以及另外两群推定的骨髓来源巨噬细胞 (BMD):BMD TAM 1(F13A1、TMEM163和MS4A4E)和 BMD TAM 2(TGFBI、MALT1和RGS2)。小胶质细胞主要分为活化前小胶质细胞(CCL3、EGR3和NFKBID)和稳态小胶质细胞(P2RY12和TMEM119)(图4A-C),这与在人和小鼠中观察到的分类一致。与先前在成人胶质母细胞瘤中的观察结果一致,在整个队列中,治疗后髓系细胞群总体上从小胶质细胞转变为骨髓来源的巨噬细胞(图4D-E)。在 IDH/H3-WT 患者的亚组分析中,他们发现所有 IDH/H3-WT 患者的髓系细胞中活化前小胶质细胞均减少。鉴于髓系细胞群的变化在不同患者间差异很大,他们应用广义线性混合模型方法来鉴定肿瘤进展过程中假性髓系细胞的通路水平变化。他们观察到干扰素和炎症反应通路上调,而与增殖和细胞代谢相关的通路(例如氧化磷酸化和E2F靶点)下调(图4F)。
既往研究发现,胶质瘤相关巨噬细胞与肿瘤细胞状态存在差异性相互作用,从而改变其活性和分化状态。事实上,他们在63个snRNA-seq数据集(大多数样本包含两个区域)中观察到肿瘤细胞状态与髓系亚型之间的频率相关性存在差异。MES样状态与促血管生成的TAM相关,而OPC/NPC样状态与稳态小胶质细胞相关(图4G)。因此,他们旨在通过分析推断的配体-受体相互作用(图4H),阐明这些pHGG相关髓系亚群如何与新定义的pHGG特异性肿瘤细胞状态相互作用。他们观察到广泛的双向细胞相互作用。重要的是,髓系细胞表达的基因有可能通过直接接触和分泌因子介导多种肿瘤细胞功能。这些相互作用包括参与调控生长和增殖(例如,SPP1-CD44 和 HBEGF-EGFR/ERBB2)、细胞黏附和迁移(例如,FN1-ITGA3/ITGB1)以及电化学或突触特性调节(例如,CALM1-KCNQ5)的过程。AC 样细胞和过度2的细胞状态预计能够最广泛地接收来自髓系亚型的细胞间信号,其受体包括 FGFR1、IGF1R 和 CD44。CD44已知能与多种配体(例如,HBEGF、PSEN1、SPP1和 VEGFA)相互作用,并在成人胶质瘤中发挥关键作用。相比之下,GPC 样细胞和 Transition 1 细胞群是最惰性的肿瘤细胞状态(图4H)。使用 CellChat将单个相互作用汇总后,在通路水平上验证了这些结果。最后,他们利用空间转录组学数据验证了髓系细胞与肿瘤细胞配体-受体相互作用在多个关键通路(包括FGF、BMP、TGFβ、NOTCH和CD44信号通路)中空间邻近细胞中富集(图4I-J)。综上所述,该分析全面表征了pHGG的髓系亚型,并表明TAM群体与肿瘤细胞状态存在差异性共存。
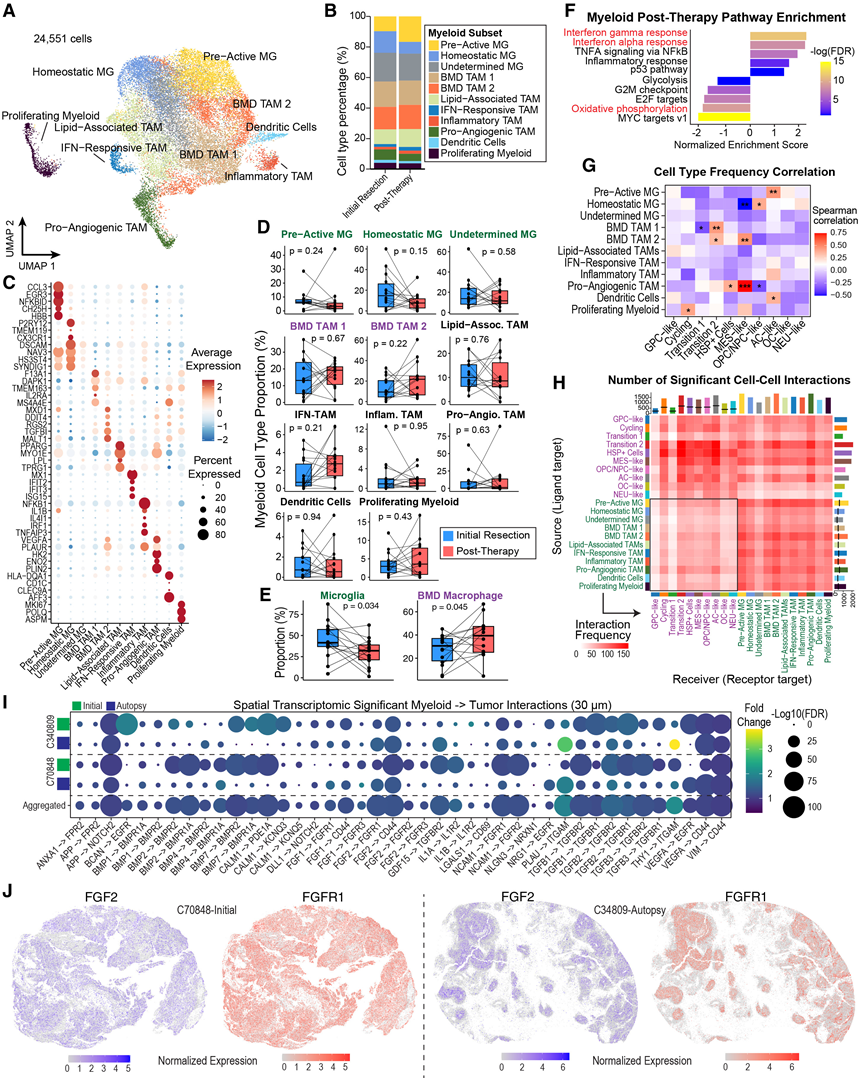
图4. 髓系对疾病进展和治疗的反应。
(A) 整合纵向pHGG snRNA-seq图谱中鉴定的已注释肿瘤相关髓系细胞群的UMAP投影。(B) 堆叠条形图,比较初始切除和治疗后样本中髓系细胞类型的组成。(C) snRNA-seq 数据中髓系细胞群代表性基因的表达。(D) 每位患者在初次切除术和治疗后时间点之间髓系细胞类型比例的变化。(E) 与 (D) 相同,在 snRNA-seq 中,初次切除和治疗后时间点之间,小胶质细胞类型比例(激活前、稳态和未定型)以及骨髓来源的 TAM 比例(BMD TAM 1 和 BMD TAM 2)发生了变化。(F) 对髓系细胞内的标志性通路进行基因集富集分析(GSEA)。(G) snRNA-seq数据集中肿瘤细胞状态比例与髓系细胞比例之间的Spearman相关系数热图。(H) 热图显示了数据集中髓系细胞和肿瘤细胞群之间显著相互作用的数量。(I) Xenium空间转录组学数据中显著的髓系细胞与肿瘤细胞间的相互作用。(J) 两个具有代表性的 Xenium 样品显示FGF2和FGFR1的空间共定位。
05
绘制pHGG的空间分布图
成人胶质瘤中肿瘤细胞和免疫细胞群复杂的拓扑定位形成了具有不同分子功能和治疗脆弱性的空间微环境。为了表征HGG的空间分布,他们采用基于索引的共检测(CODEX)空间蛋白质组学方法,利用包含52个抗体的panel,对11个具有配对snRNA-seq数据的全切片福尔马林固定石蜡包埋(FFPE)样本进行了分析,其中包括3对IDH/H3-WT患者匹配的纵向样本(图5A)。在确认抗体染色形态和共定位结果正确后,他们证实CODEX panel能够分辨包括肿瘤主体、灰质和白质在内的宏观解剖结构(图5B)。随后,在对单个细胞进行分割、整合和聚类后,他们对超过750万个单个细胞进行了注释(图5C)。他们捕捉到了肿瘤细胞状态的主要轴,从前神经细胞(SOX2、OLIG1和OLIG2)到星形胶质细胞/间质细胞(CD44、VIM和GFAP)。他们还观察到一种高表达APOE和SPP1的肿瘤细胞群,该细胞群主要存在于坏死周围区域。肿瘤细胞状态在不同样本之间以及同一样本内部均呈异质性分布,肿瘤的某些区域主要由前神经细胞或间质细胞斑块组成(图5D-E)。
髓系细胞群呈现出表型分布,包括小胶质细胞和巨噬细胞(图5E)。他们鉴定出一个高表达CD163和CD206的巨噬细胞群,另一个以高HLA-DR表达为特征的巨噬细胞群,以及大量形态异质的MPO+髓系细胞,这些细胞可能代表中性粒细胞和MPO+单核细胞,正如先前报道。该MPO+细胞群高表达HIF-1α,并且在多个样本中主要以坏死周围区域的大量浸润形式存在。虽然他们鉴定出少量CD4+和CD8+ T细胞,但图像检查显示,T细胞主要位于血管内或集中在出血区域。这表明组织中的血液污染物可能会干扰对罕见免疫细胞群的单细胞测序分析。最后,他们观察到免疫检查点分子[包括 CD47 和程序性死亡配体 1 (PD-L1)]的表达具有空间限制性。
为了系统地鉴定所有样本中反复出现的空间模式,他们首先进行了无监督邻域分析,并根据细胞类型的相对富集程度,识别出15个细胞邻域(CN),并对其进行了手动标注(图5F)。这些邻域在样本中呈异质性分布,并涵盖了预期的解剖结构,包括灰质(CN2,成熟神经元)、白质(CN6,成熟少突胶质细胞)和浸润性肿瘤区域(CN7,少突胶质细胞和肿瘤细胞)(图5G)。此外,该分析还突出了以不同肿瘤细胞状态(即前神经元、中间型和间质细胞邻域)为主的局部区域,以及MPO+浸润和血管邻域(图5F)。肿瘤细胞倾向于与具有相同表型的细胞共定位,例如前神经元肿瘤细胞与其他前神经元肿瘤细胞共定位(图5D-F)。值得注意的是,间质肿瘤细胞类型在任何细胞邻域中均未与神经前体肿瘤细胞共富集,无论是在总体上还是按亚型分别计算的邻域中(图5F)。免疫细胞在不同的细胞邻域中存在差异性分布。小胶质细胞主要富集于正常脑组织区域,尤其是灰质(图5F),并且观察到它们集中在肿瘤-正常组织交界处。巨噬细胞和T细胞共同富集于免疫细胞富集邻域(CN1)、血管周围邻域(CN5)、血管肿瘤邻域(CN11)、间质肿瘤邻域(CN3)以及肿瘤/免疫细胞混合邻域(CN9)。在所有神经前体肿瘤细胞显著富集的邻域中,免疫细胞相对减少(图5F)。通过检测肿瘤细胞与髓系细胞的距离,他们发现神经前体和间质肿瘤细胞最常富集于HLA高表达巨噬细胞附近,而坏死周围肿瘤细胞则与MPO阳性髓系细胞和未分类巨噬细胞共定位。此外,他们发现间质肿瘤细胞距离内皮细胞最近,而坏死周围肿瘤细胞距离血管最远。
接下来,他们试图鉴定治疗后富集的空间模式。为了减轻样本间显著的异质性,他们分别对每个样本进行邻域划分,并利用近期开发的N-Orbit框架,通过主成分分析对每个相邻邻域实例进行投影。他们发现第一主成分(PC1)捕捉到了时间点效应(图5H)。然后,他们识别出两个对PC1贡献最大的聚类,这两个聚类包含的子聚类在初始切除或治疗后时间点富集(图5I-J)。每个聚类都包含了这些时间点所有患者的样本。将治疗后聚类1中的邻域实例追溯到其富集的N-Orbit,结果显示少突胶质细胞富集,这些少突胶质细胞主要与前神经元和中间型肿瘤细胞相互作用,并伴有涉及小胶质细胞等免疫细胞的次要相互作用(图5K-L)。聚类2主要捕捉了血管特征。这些结果表明,治疗后与少突胶质细胞的空间协调性增强,并且这些相互作用有利于神经前体和中间型肿瘤细胞表型,而非间质型肿瘤细胞表型。
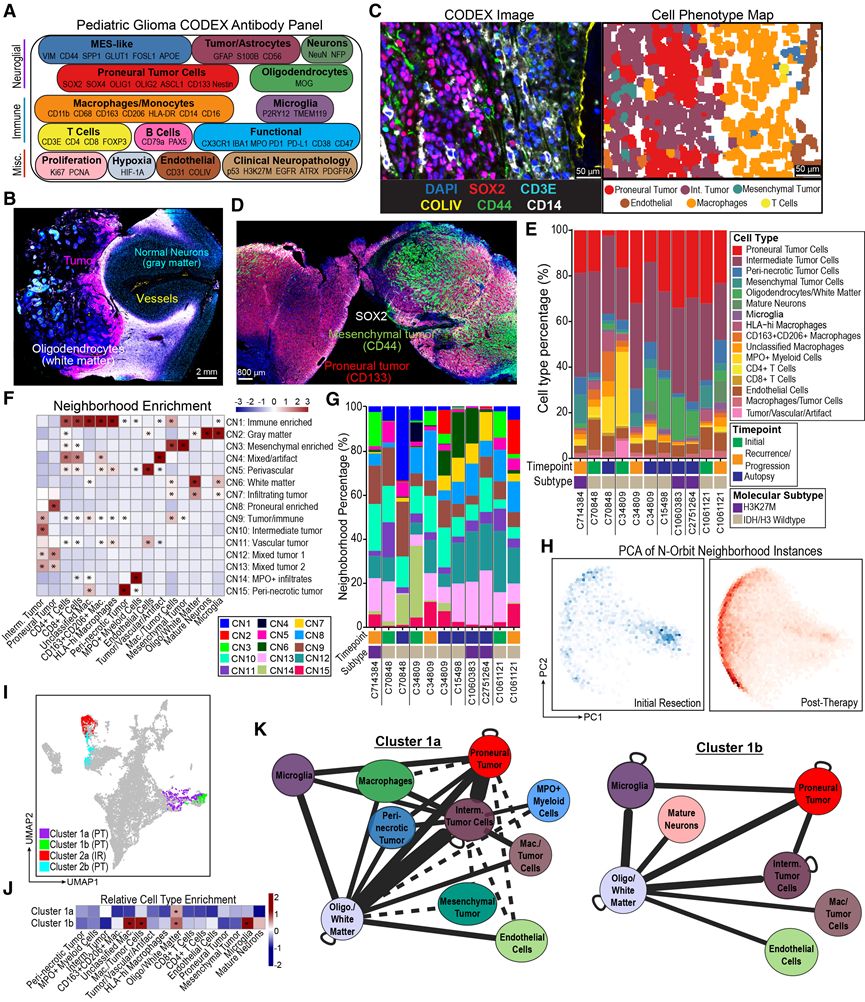
图5. CODEX 成像揭示了 pHGG 治疗过程中微环境的变化。
(A) 图示抗体 CODEX组合按靶细胞群或细胞功能划分。(B) 代表性的 CODEX 图像,突出显示肿瘤肿块和正常脑组织的亚结构。(C) 带有选定荧光标记的 CODEX 图像(左)与细胞表型图(右)配对。 (D) 代表性的CODEX图像展示了空间分布受限的肿瘤细胞状态群体。(E) 每个 CODEX 样本中的细胞类型比例。(F) 热图显示了各亚群中细胞类型的相对富集情况。(G) 每个 CODEX 样本中的邻里比例。(H) N-Orbit 导出的邻域距离矩阵的主成分分析密度图。(I) 在 UMAP 投影上表示对 PC1 贡献最大的邻域聚类。(J) 聚类1a 和 1b 的细胞类型富集。(K) 治疗后富集邻域实例聚类 1a 和 1b 的汇总子图。
06
肿瘤亚克隆动态揭示反复出现的基因组改变
接下来,他们旨在利用纵向数据来识别疾病进展和治疗过程中基因组和转录组的变化。他们首先利用大规模拷贝数变异(CNV)数据,通过 Clonalscope 追踪患者匹配样本中的肿瘤亚克隆。Clonalscope 整合了snRNA-seq和匹配的全基因组测序(WGS)数据。他们定义了每位患者最早时间点的肿瘤亚克隆,并追踪至后续治疗时间点,以评估扩增或消退的细胞群。通过这种方法,他们在14例患者中识别出了谱系可追溯的肿瘤亚克隆,每位患者包含4至9个亚克隆,且不同时间点的克隆动态存在差异(图6A)。他们鉴定出一些复发性CNV,包括1q、7p/7q、8q、19p/19q和20p染色体上的拷贝数增加,以及5p/5q、6q、10p/10q和14q染色体上的拷贝数缺失(图6B-C)。对肿瘤亚克隆中基因水平的CNV进行聚类分析,揭示了患者间高度相关的CNV复发模块,表明在疾病进展过程中染色体改变的动态模式相似(图6D)。值得注意的是,某些改变在扩增的亚克隆中更频繁地发生(例如,18p和19q染色体上的拷贝数增加以及14q染色体上的拷贝数缺失),提示这些改变可能赋予生存优势(图6E)。然而,他们发现较大的基因组/亚克隆变化与肿瘤细胞状态组成的较大变化并不相关(图6F)。最后,他们将扩增的亚克隆与每位患者剩余的恶性细胞进行比较。结果显示,大多数患者的免疫相关通路均上调,表明肿瘤表型的选择可能与肿瘤微环境有关(图 6G)。
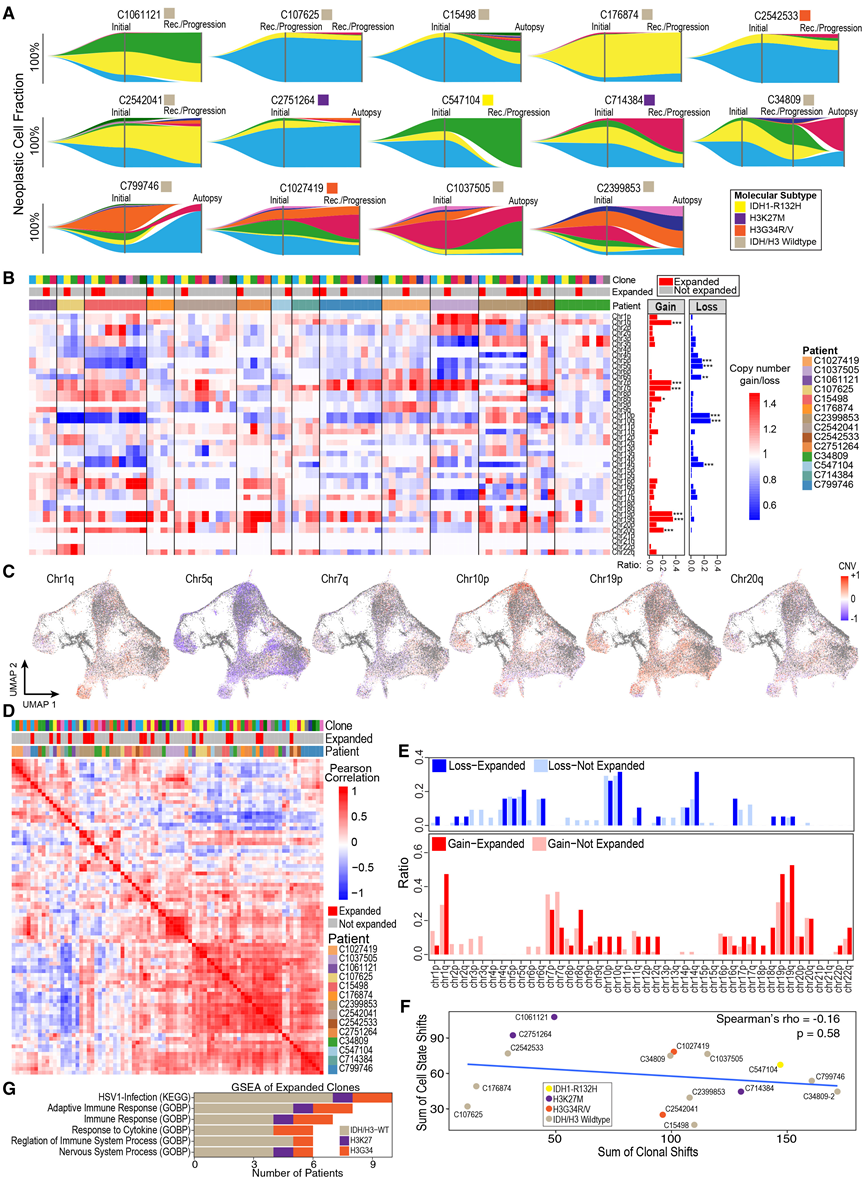
图6. 患者间肿瘤亚克隆动态变化。
(A) 鱼尾图显示了 14 名患者肿瘤亚克隆群体在纵向时间点的变化。(B) 左图:使用Clonalscope 分析14例患者中每个肿瘤亚克隆在染色体臂水平上的平均 CNV 热图。克隆颜色(顶行)对应于(A)中所示的患者特异性亚克隆。右图:每个染色体臂的二值化拷贝数增加或减少比率。(C) UMAP 投影显示肿瘤细胞,颜色根据 Clonalscope 预测的特定染色体臂片段的扩增或缺失情况而定。(D) 基于基因水平拷贝数变异(CNV)估计值的亚克隆相关性热图。(E) 染色体臂水平上二元化拷贝数变异(拷贝数增加或缺失)的比率,比较扩增克隆和非扩增克隆。(F) 亚克隆和细胞状态转变之间的关联性。(G) 在扩增的亚克隆中富集特定通路的患者数量。
07
纵向分析揭示肿瘤细胞内在靶点
随后,他们采用类似的广义线性混合模型(GLMM)方法,识别在所有肿瘤细胞中,于治疗时间点上调的基因和通路,并考虑个体患者差异。尽管未观察到肿瘤细胞群水平的细胞状态转变,但该分析仍得出627个显著上调的基因和1551个显著下调的基因(图7A)。通路水平的变化分析显示,I型和II型干扰素反应通路以及神经活性配体-受体相互作用基因集上调,而与细胞增殖和代谢相关的通路(主要是氧化磷酸化)下调(图7B)。他们推测,辐射效应可能与观察到的转录变化存在因果关系。事实上,他们采用类似的广义线性混合模型(GLMM)方法分析了三种未经治疗的pHGG细胞系在治疗前后的RNA-seq数据,发现电离辐射治疗后,这些细胞系也出现了许多相同的通路变化(图7C-D)。这表明放射治疗的直接作用在于抑制细胞周期和氧化磷酸化,并增强干扰素和神经活性信号传导。
随后,他们旨在利用这种肿瘤细胞特异性纵向分析,在pHGG细胞系中鉴定肿瘤细胞内在的药物靶点。为了确定基因靶点的优先级,他们筛选了在肿瘤进展过程中差异上调的基因,并将其与多个药物靶点数据库和癌症依赖性图谱(DepMap)进行比对,同时考虑了它们在肿瘤微环境中作为受体的作用(图7E)。在筛选出需要验证的靶点后,他们筛选了24种药理化合物,并在6种pHGG细胞系中评估了它们在72小时内对细胞增殖和活力的影响。对这些细胞系的批量RNA测序数据进行反卷积分析表明,它们重现了在患者样本中观察到的肿瘤细胞状态谱。他们验证了AZD4547(FGFR抑制剂)、danusertib(Aurora激酶/ABL激酶/FGFR1抑制剂)、panobinostat(非选择性HDAC抑制剂)和trametinib(MEK抑制剂)对多种细胞系的细胞毒性作用(图7F)。治疗进展后,多种与细胞凋亡和炎症小体激活相关的基因表达上调,包括caspase(CASP1、CASP4)、BCL2L1和BCL6。事实上,与DMSO对照组相比,使用belnacasan抑制CASP1以及使用navitoclax或venetoclax抑制BCL-2均可降低体外细胞增殖。值得注意的是,曲美替尼与纳维托克在多种细胞系中表现出协同作用,这表明凋亡抑制剂可以改善现有疗法的疗效(图7G)。与通路分析一致,他们观察到参与电化学和突触通讯的多个基因显著上调,这些基因已证明能够促进胶质瘤的进展和侵袭。选择性CHRM3拮抗剂J-104129通过诱导细胞凋亡导致显著的细胞死亡(图7D)。选择性GABA A受体拮抗剂加巴嗪在多种细胞系中表现出轻微的抗增殖作用。最后,他们的筛选结果显示,分泌型磷脂酶A2(sPLA2)值得在儿童胶质瘤中进行进一步研究,在所有六种细胞系中,使用 PLA2 抑制剂 varespladib 治疗后均表现出剂量依赖性反应,并且发现其作用机制是通过诱导细胞凋亡(图 7 D)。
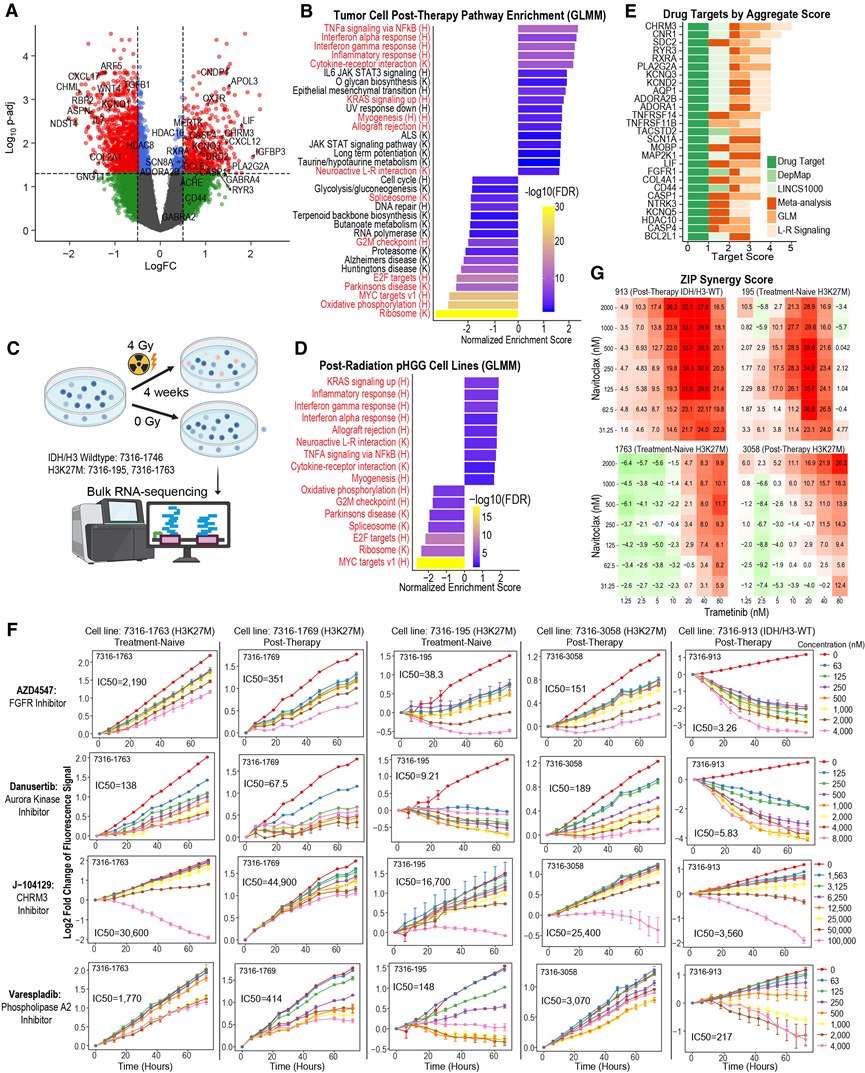
图7. 通过体外药物筛选鉴定治疗敏感性。
(A) 采用线性混合模型识别肿瘤细胞在初次切除和治疗后时间点之间的差异表达基因。(B)对中所有基因的 Hallmark和KEGG通路进行GSEA。(C) 辐射实验示意图。(D) 使用与中类似的广义线性混合模型 (GLMM) 对三种 pHGG 细胞系进行辐射后变化的GSEA。(E) 按综合排名得分排名的前几名基因靶点。(F) 体外药物筛选中,在球状体培养条件下培养的人类pHGG细胞系的生长曲线。(G) 曲美替尼和纳维托克组合在不同浓度下的协同作用评分。
+ + + + + + + + + + +
结 论
本研究通过多组学测序对16例pHGG患者的分子异质性队列进行了纵向分析,以捕捉疾病进展和治疗过程中肿瘤细胞和微环境特征的演变,定义了一组核心的pHGG肿瘤细胞状态,并观察到恶性细胞表型之间存在不同的肿瘤-髓系相互作用。治疗后关键神经调节因子和干扰素反应上调,表明它们是恶性细胞的内在靶点。本研究观察到少突胶质细胞在疾病进展过程中增加,并且它们与促神经肿瘤细胞协调空间结构。该纵向 pHGG 多组学图谱捕捉了治疗反应的特征,并为儿科脑肿瘤的研究提供了新的理论依据。
+ + + + +



