

 English
English文献解读|Nat Commun(15.7):通过对巴马小型猪的转录组学和蛋白质组学分析揭示围产期脑发育转变
✦ +
+
论文ID
原名:Perinatal brain developmental transition revealed by transcriptomic and proteomic analyses of Bama miniature pigs
译名:通过对巴马小型猪的转录组学和蛋白质组学分析揭示围产期脑发育转变
期刊:Nature Communications
影响因子:15.7
发表时间:2026.04.02
DOI号:10.1038/s41467-026-71360-9
背 景
围产期,即从妊娠晚期到婴儿早期,是神经发育的关键阶段,其特征是脑总量较妊娠中期增加三倍,皮质表面积增加四倍。在这个关键时期,发育的重点从神经元生成转向神经回路的精细化,同时胶质细胞生成进入高峰期,表现为星形胶质细胞树突化加速和少突胶质细胞介导的髓鞘形成。这些过程在自闭症谱系障碍(ASD)、小头畸形和无脑回畸形等神经发育障碍中均存在缺陷。因此,理解围产期的分子动力学对于解析正常的神经发育和病理生理级联反应至关重要。尽管围产期具有重要的研究意义,但由于获取完整组织样本存在诸多伦理障碍,围产期仍然是人类大脑发育生物学中研究最少的阶段。因此,调控围产期神经发育的机制,包括转录组转换、蛋白质组成熟和多组学相互作用,在很大程度上仍不为人知。
为了弥补这一关键空白,研究团队利用了人类和巴马小型猪在神经发育上的趋同性,因为这两个物种都经历了从妊娠晚期到出生后阶段的持续性围产期脑扩张。这克服了无脑回啮齿动物的根本局限性,它们的妊娠期较短(19-21天),缺乏相应的围产期过渡。此外,仔猪能够重现人类婴儿神经解剖结构的关键特征,包括脑沟-脑回结构、灰质和白质分布以及脑重比。
实验设计
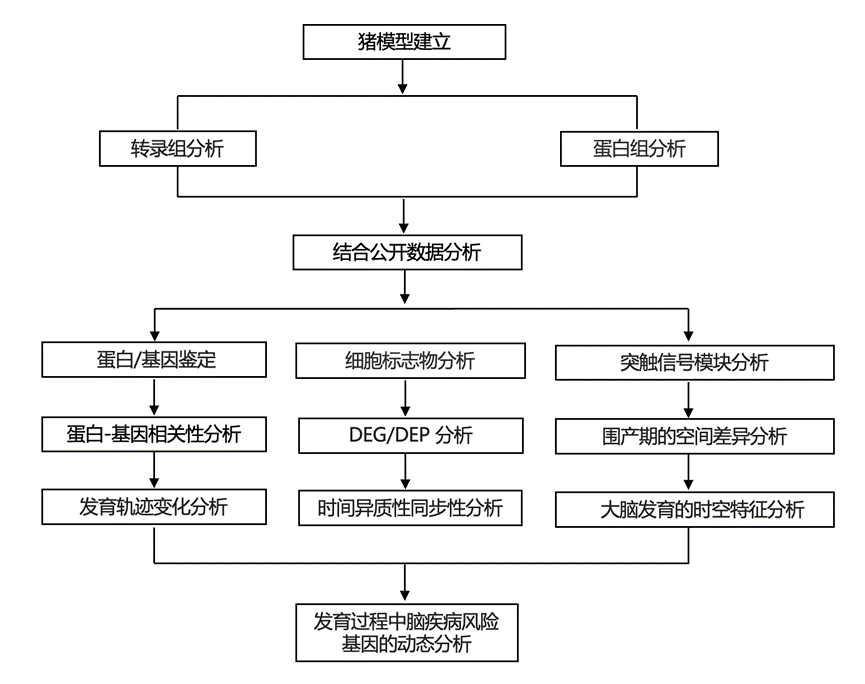
结 果
01

巴马小型猪脑发育过程中转录组和蛋白质组的时空概览
研究团队对从多个脑区采集的336个样本进行了转录组分析(RNA-seq)和蛋白组分析,这些脑区包括前额叶皮层(PFC)、颞叶皮层(TMP)、枕叶皮层(OCC)、纹状体(STR)、丘脑(THA)、海马(HIP)和小脑(CERE)。这些样本来自30头胎猪和仔猪,每个样本的转录组和蛋白质组大多来源于同一组织,仅有6个样本(占168个样本的3.57%)来自同一脑区内的不同个体(图 1a)。所有样本均采集于八个主要发育阶段:五个来自妊娠晚期(胚胎第76、85、94、104和109天),三个来自出生后(出生后第0、3和30天)。每个时间点均包含至少两个生物学重复(P0 时的 CERE 除外)(图 1a),涵盖了神经发生、胶质细胞发生、突触发生、髓鞘形成、皮层扩张、神经回路建立以及最终形成功能性脑网络等关键发育阶段。在此期间,体重和脑重均迅速增加,但增长轨迹截然不同。脑重在妊娠晚期持续增长,与妊娠晚期早期(E76)相比,出生时脑重增加了五倍,这与围产期观察到的特征性皮层扩张相一致。相比之下,体重在出生后迅速增加,脑重与体重的比值在 E94 附近达到峰值,然后持续下降,这表明猪的生长发育最初优先考虑脑部扩张,而在围产期则转向更注重全身发育的模式。未观察到基于性别的体重、脑重或脑重与体重比存在统计学上的显著差异。
经过严格的质量控制,他们在每个年龄组至少两个样本中检测到约30000个基因和8000个蛋白质,其中超过90%的已鉴定蛋白质在转录组中检测到了其对应的基因(图 1b)。在所有已鉴定的基因和蛋白质中,约有10000个基因和6,000个蛋白质的表达水平超过总表达量的一半,超过80%的蛋白质可以与其对应的基因匹配(图 1b)。主成分分析(PCA)显示时间进程是主要的生物学变量,转录组和蛋白质组数据集均沿PC1按时间顺序排列(分别占总方差的24.81%和24.93%)(图 1c-d),这与胚胎发育模式一致。沿PC2的空间组织呈现出皮层(PFC/TMP/OCC)-皮层下(STR/THA/HIP)-小脑(CERE)的层级结构,在转录组中更为显著(图 1c,d)。有趣的是,皮层下结构的转录异质性高于皮层区域(图 1c),而HIP则聚集在新皮层附近,这可能反映了异皮层和新皮层的进化同源性及其相似的细胞组成(图 1c)。小脑样本显示出受限的发育轨迹和跨组学层级的明显空间分离(图 1c-d),证实了其独特的成熟时间线。这些关键的时空模式在非线性降维分析(例如 UMAP和 t-SNE)中得到了一致的重现。跨组学整合显示出显著的时间相关性和空间相关性,前 20 个主成分的一致性达到 60%(图 1e)。综上所述,他们构建了巴马小型猪脑多个区域和发育时间点的转录组和蛋白质组综合图谱,揭示了其发育过程中独特的时空特征。该资源已适配在线交互式探索,可通过http://www.braininfo.ac.cn/PBAtlas/免费访问。
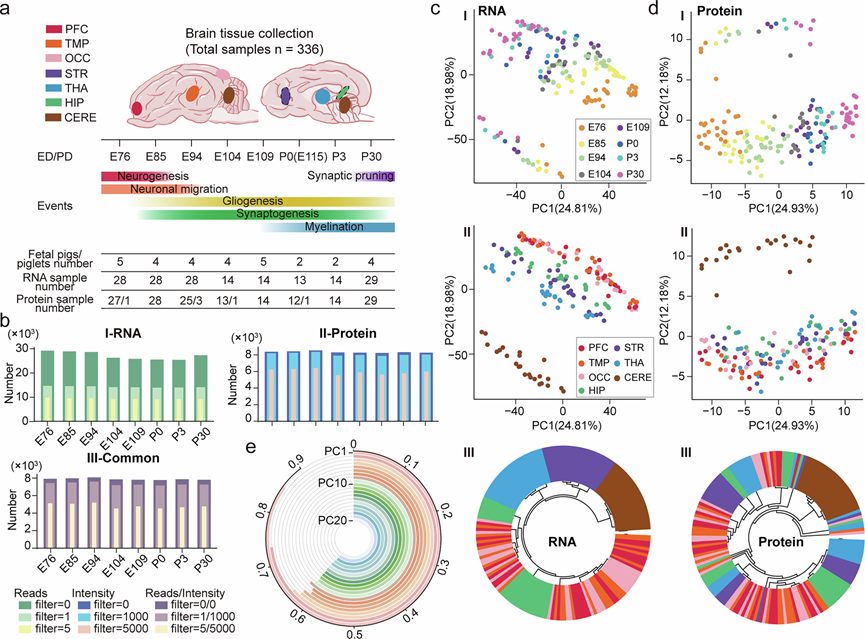
图1. 巴马小型猪脑发育过程中转录组和蛋白质组分析的概述。
(a) 巴马小型猪脑组织样本采集示意图。(b) 基因(I)和蛋白质(II)的数量及其重叠部分(III)。(c-d) 转录组和蛋白质组数据中按年龄和区域分组的样本的主成分分析(PCA)图。(e) 转录组和蛋白质组数据前 20 个主成分之间的相关系数。
02
模型重现了灵长类动物围产期神经发育
为了研究大脑发育过程中进化保守的转录程序,他们将猪皮层和小脑RNA测序数据与已发表的多物种发育转录组数据集(包括人、恒河猴、小鼠、大鼠、兔、负鼠和鸡)进行了跨物种整合,该数据集涵盖了从产前到成年的阶段。主成分分析(PCA)显示,皮层(CTX)和小脑(CERE)样本均沿第二主成分(PC2)呈现物种分离。值得注意的是,人、恒河猴和猪的样本形成了一个紧密聚集的组,与啮齿类动物(小鼠/大鼠/兔)和原始脊椎动物(负鼠/鸡)明显不同,这与Cardoso-Moreira等人提出的进化层级相吻合(图 2a-b)。这种保守的系统发育模式得到了层次聚类的证实,其中三个脑回物种(人类、恒河猴、猪)与无脑回生物相比表现出更优异的转录相似性(图 2c)。
发育阶段是PC1轴上的主要生物学变量,所有物种的样本均按时间顺序排列,从产前阶段到成年阶段(图 2a-b)。PC1轴捕捉到了物种间的异时性,其中人类的发育轨迹最长(PC1跨度最大),而鸡的发育轨迹最短,这与已知的神经发育时间线一致(图 2a-b)。至关重要的是,猪和恒河猴的样本在PC1轴上占据了过渡位置,填补了人类围产期覆盖范围的关键空白(图 2a-b)。当应用RAPToR的跨物种年龄映射算法时,这种时间互补性尤为明显。该算法显示,猪的围产期皮层样本与人类的产前晚期到产后早期阶段完全吻合(图 2d)。具体而言,猪胚胎发育第76天至第104天(E76–E104)对应于人类受孕后第183天至第257天(PCD 183–257),准确地将其定位在人类围产期窗口(PCD 182–273)内。虽然大鼠、兔、负鼠和鸡等物种在围产期也有一些代表,但与猪相比,它们的时间跨度仍然明显不足,这凸显了猪围产期图谱的独特价值。至于猪小脑样本(图 2d),除两个来自出生后第30天(P30)的样本(S6-3-LR-Cere和S12-1-Cere,占8.33%)外,其余所有样本均落在人类围产期范围内。这表明小脑发育窗口相对于皮层而言较为狭窄,年龄匹配的CERE样本与CTX样本中PC1跨度较短也证实了这一点(图 2d)。这一发现与先前在猴子中的报道一致,并暗示哺乳动物的神经发育序列是保守的(图 2d)。
为了从分子层面验证猪模型模拟人类脑围产期发育的能力,他们对人类全生命周期和猪围产期转录组进行了跨物种整合分析。该分析揭示了人类和猪之间平滑的过渡,包括以少突胶质细胞特异性标记物(MBP、MOG、PLP1)和星形胶质细胞特异性标记物(GFAP、AQP4)为标志的胶质细胞生成程序的围产期激活,这些分子特征通常与灵长类动物脑成熟后期相关(图2e)。值得注意的是,他们观察到围产期小白蛋白(PVALB)表达的启动,PVALB是一种钙结合蛋白,可定义GABA能中间神经元的身份,这表明PVALB+中间神经元的分子特化始于这一过渡时期,与脑回物种中保守的发育机制一致(图 2e)。此外,他们还发现神经调控基因AK5(腺苷酸激酶5)和CST3 (胱抑素C)在围产期特异性激活,这些基因在人类妊娠早期/中期数据集中转录沉默,但在出生后持续表达,从而填补了灵长类动物围产期转录组动态变化中的关键知识空白(图 2e)。值得注意的是,与上述基因激活模式相反,他们还发现了一种反向沉默模式:在人类中表现为早期产前表达后产后抑制的基因(例如ISLR2、TUBB2)在猪围产期表现出逐渐下调(图 2e)。此外,他们在已发表的人类单细胞数据集中验证了这些基因的表达模式。在人类和猪的 CTX 中进行的免疫荧光验证表明,AK5 和 PVALB 在妊娠第 23 周 (GW23) 时均未表达,而 AK5 的表达在早期围产期(猪为 E85)开始,PVALB 的表达在晚期围产期(猪为 E109)开始,从而在时间上连接了人类的产前沉默和产后激活(图 2f)。除了单基因分析外,他们还评估了围产期相关的生物学通路,发现猪的发育轨迹与人类参考序列高度吻合,大多数样本与拟合曲线紧密对齐,并准确定位在人类围产期窗口内。这些整合的分子分析表明,猪可以作为研究灵长类动物出生过渡期转录调控的动态模型。蛋白质组学整合进一步扩展了这些发现:尽管蛋白质组范围内的保守性限制了PC2上的物种水平分辨率,但猪脑蛋白表达谱在食蟹猴的发育轨迹中占据了E90-P3的连续谱(图 2g)。除了分子转变之外,他们还在关键生物学事件中发现了类似的连续谱,例如弓状核(ARC)中间神经元的迁移流。虽然之前的研究报道了这种迁徙流在出生后的存在,但研究结果表明,它在出生前就已出现,并在出生后持续存在,这一模式与人类的观察结果一致。总而言之,这种多层次的一致性表明,猪是研究啮齿动物模型无法触及的灵长类特有围产期过程的合适模型,填补了灵长类围产期研究的空白,并提供了多层次的生物学证据,证明其能够通过进化保守的机制重现人类特有的发育过程。
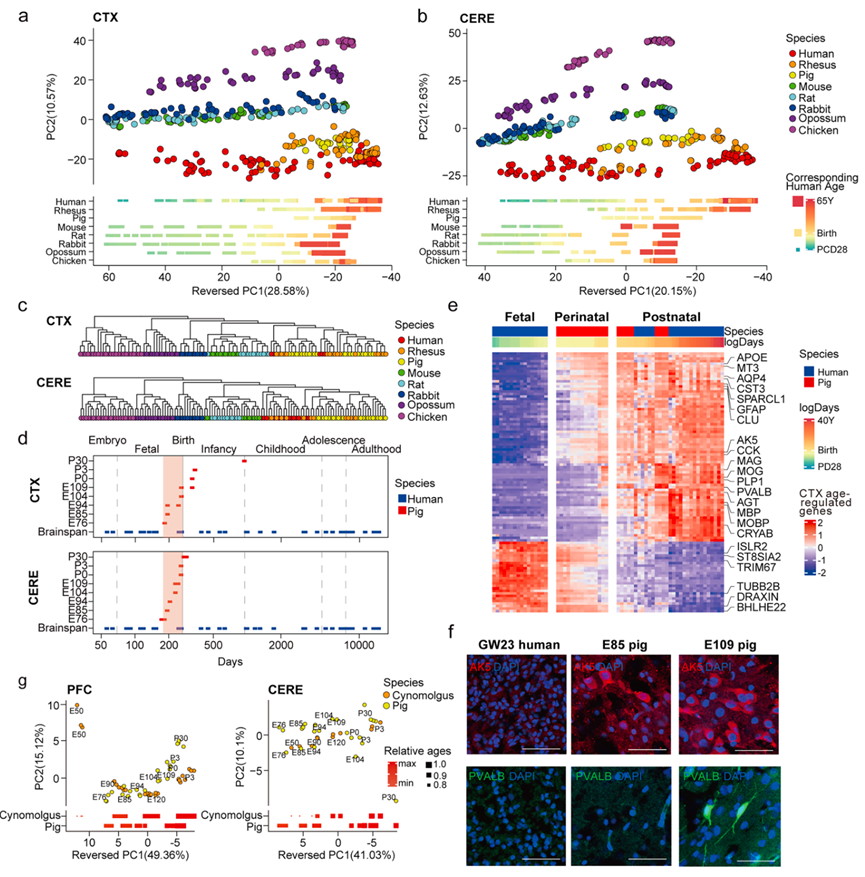
图2. 皮层和小脑发育的跨物种转录组和蛋白质组整合。
(a-b) 基于跨物种转录组整合的皮层和小脑样本的主成分分析(PCA)。(c) 基于转录组相似性,使用平均连接法对多物种整合数据集进行层次聚类,分别对皮层(上图)和小脑(下图)样本进行分析。(d) RAPToR算法预测的猪皮层(上图)和小脑(下图)样本的人类等效发育年龄。(e) 通过整合分析人类寿命和猪围产期转录组,鉴定出皮层发育中年龄调控基因的热图。(f) 免疫荧光分析。
03
围产期发育过程中观察到的重大转变
基于区域的转录组和蛋白质组主成分分析(PCA)揭示了沿第一主成分(PC1)的保守V形时间轨迹,其中E76-E94(对应于人类PCD 183-206)和E104-P30(对应于人类PCD 257-出生后673天)阶段之间存在明显的转折点,在转录组PFC/OCC/THA/HIP样本和蛋白质组PFC/OCC/THA样本中尤为显著(图 3a-c)。区域相关性分析支持了这些发现,揭示了E94和E104之间的最大差异,进一步强化了PCA中观察到的时间转变(图 3b-d)。值得注意的是,这些围产期中期转变与人类晚期产前发生的保守发育事件相对应。然而,他们对猪的多组学分析获得了更高的时间分辨率,从而建立了一个进化上保守的模型,该模型精确地描绘了这一关键的神经发育转变。
为了从分子层面解析发育转变,他们系统地分析了跨越皮层和皮层下区域的七个连续年龄阶段的差异表达基因和蛋白质(DEG/DEP)。在E94-E104转变期间,所有脑区均观察到DEG显著增加,虽然OCC在峰值后呈现更为缓慢的下降趋势(图 3e-f)。相比之下,DEP的区域异质性更为明显。虽然E94-E104转变期相似,但某些区域的DEP转折点出现延迟或波动,OCC甚至在E109-P0时形成第二个峰值(图 3f)。转录和翻译调控之间的这种不同步可能源于蛋白质组学固有的技术限制,例如在特定发育时间点蛋白质覆盖率较低或取样偏差(图 1b)。
他们将时间调控基因/蛋白(TRG/TRP)定义为仅在单一发育时间窗口内出现的差异表达基因/蛋白(DEG/DEP),其中超过80%的TRG/TRP在大多数脑区于E94-E104达到峰值(图3g-h)。值得注意的是,TRP在转变后仍保持高表达水平,表明功能成熟过程中蛋白质组持续重塑(图 3h)。对皮层、皮层下和小脑样本中TRG和TRP的比较富集分析揭示了关键发育转变过程中区域特异性的分子通路激活动态(图 3i-j)。皮层区域表现出顺序激活模式,早期富集核心细胞过程(RhoA信号通路、TGF-β通路)和轴突发生相关通路(SLIT-ROBO介导的轴突导向),后期发展至高级突触调控(红藻氨酸受体突触前功能、GABA能信号通路)。皮层下区域最初激活轴突导向机制(Slit-Robo介导的连合轴突导向)和神经元迁移生物过程(LIS1通路),随后过渡到凋亡调控和脂质代谢过程,这些过程可能促进神经回路的精细化。值得注意的是,小脑表现出独特的发育层级,优先进行早期胶质细胞生成和突触形成,之后才过渡到回路精细化。
最后,他们鉴定出一组在胚胎第104天(E104)表达显著升高的基因/蛋白(图 3k),其中髓鞘碱性蛋白(MBP)在所有检测的脑区均呈泛区域性上调,这一模式已通过定量实时PCR(qPCR)和免疫荧光分析得到验证(图 3l-m)。这种同步的MBP诱导表明少突胶质细胞生成旺盛,与髓鞘形成高峰期相吻合,提示神经网络中存在大规模的突触整合,这一发现与他们之前的通路分析结果一致,该分析揭示了突触信号模块的富集(图 3i-j)。此外,他们发现一些区域特异性基因在此期间表达激增,包括运动协调相关基因KRT25、皮层下产热调节因子TMEM26和小脑炎症介质LCN2,这些基因均通过qPCR验证(图 3l)。综上所述,多组学研究表明,E94-E104代表了猪神经发育过程中一个同步的发育转折点。
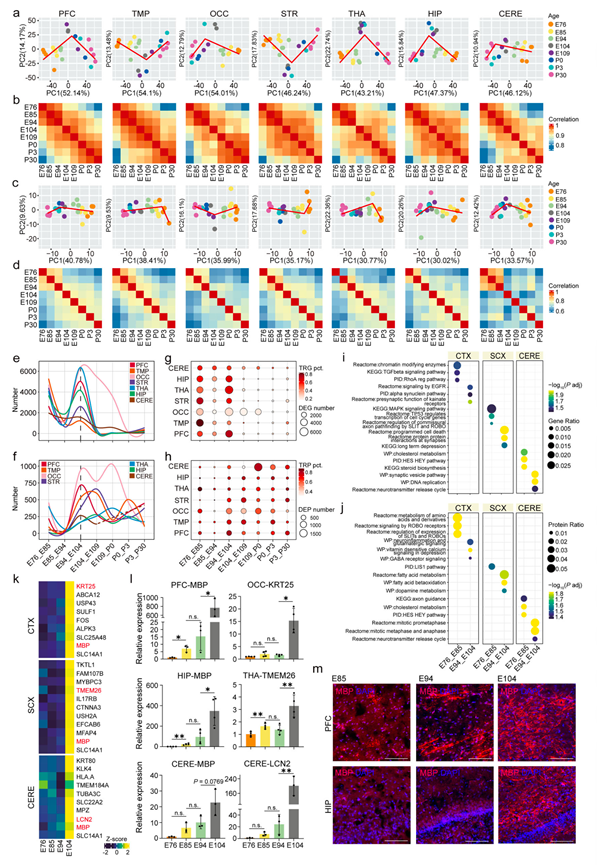
图3. 围产期大脑发育各区域的时间异质性同步。
(a, c) 转录组和蛋白质组的主成分分析 (PCA) 展示了不同发育阶段脑区样本沿 PC1 和 PC2 维度的分布。(b, d) 热图展示了各脑区内不同发育阶段样本间转录组和蛋白质组的 Spearman 相关性。(e-f) LOESS 拟合曲线展示了脑区相邻时间点间DEG和DEP丰度变化(y 轴)的时间趋势。(g-h) 气泡图展示了相邻脑区时间点间DEG和差异表达蛋白DEP中TRG和TRP的比例。(i-j) 对发育阶段E76-E85和E94-E104的皮层(CTX)、皮层下区(SCX)和小脑(CERE)的TRG和TRP进行了通路富集分析。(k) 热图显示了在皮层 (CTX)、皮层下 (SCX) 和小脑 (CERE) 中表现出 E104 特异性表达激增的基因。(l) 通过密度分析法定量 mRNA 表达。(m) E85/E94/E104猪脑矢状切片中前额叶皮层(PFC)和海马(HIP)髓鞘碱性蛋白(MBP)免疫染色代表性图像。
04
发育速度揭示了围产期空间差异
为了系统地研究大脑发育过程中的空间异质性,他们建立了“发育速度”方法,该方法通过分析基因或蛋白质的时间变化来量化区域生长速率。他们发现皮层区域表现出高度同步的发育轨迹,这由紧密聚集的发育速度拟合线所证实,表明围产期发育协调一致(图 4a-b) 。皮层下区域的发育年龄跨度明显缩短,其中小脑的跨度最小。滑动窗口分析和自举重采样验证了发育速度估计的稳健性,两者均支持发育轨迹的可靠性。他们利用表达轨迹与发育年龄之间的相关性分析,探索了与发育速度存在区域特异性关联的基因和蛋白质。约48%的DVP显示出相应的DVG,且在不同脑区呈现保守的分布模式。皮层包含最丰富的DVG/DVP库,其内部区域间DVG和DVP的分子保守性分别约为54%和42%(图 4c-d)。相比之下,皮层下区域在不同区域间的分子保守性约为 30% (DVG) 和 47% (DVP),而小脑与其他区域相比,平均分子保守性约为 40% (DVG) 和 34% (DVP)(图 4c-d)。
为了阐明 DVG 的细胞相关性,他们使用区域匹配的单细胞和单核数据对整体转录组进行了反卷积分析。由于缺乏相应发育阶段猪的高质量单细胞/核转录组数据,后续分析改用了密切匹配的人类发育阶段的数据(STR 除外,该分析采用了恒河猴数据)。鉴定出全脑保守的细胞类型,包括兴奋性神经元(ExN)、抑制性中间神经元(InN)和胶质细胞(Glia),以及区域特异性细胞群,例如海马齿状回颗粒细胞(DG)、纹状体中型棘状神经元(MSN)和小脑颗粒细胞及浦肯野细胞;这种对解剖学上受限的细胞类型的清晰解析证实了他们取样的精确性(图 4e)。胶质细胞的比例在各个脑区均呈现上升趋势,这与围产期是胶质细胞生成的高峰期相一致(图 4d)。同时,高置信度的细胞类型相关DVG在所有检测的脑区均与胶质细胞表现出显著关联(图 4e)。小脑样本与此模式有所不同,这可能反映了其早熟的发育时间线,其中胶质细胞生成高峰出现在围产期之前(图 4e)。在探索其时间变化时,检测到细胞类型相关DVG的同步上调,并在E104达到表达的突然激增,这与先前确定的发育过渡点相吻合(图 3e-f和图4f)。该细胞类型相关的DVG集合包括突触组织因子(DLGAP1、CAMK2N1、CALM1、UNC13C),以及区域特异性效应因子,例如纹状体多巴胺靶基因PPP1R1B和小脑颗粒细胞标记基因CBLN1(图 4f)。此外,他们系统地研究了高置信度细胞类型相关 DVG 的空间分布。尽管 DVG 与胶质细胞群密切相关,但区域特异性 DVG 仍占主导地位,揭示了不同脑区胶质生成程序的显著异质性(图 4g)。值得注意的是,与皮层下和小脑区域相比,皮层区域的区域间异质性显著降低,这归因于皮层区域间共有的保守的细胞类型相关 DVG 特征(图 4c-g)。
接下来,他们计算了每个发育时间点脑区之间的DEG,观察到区域间异质性在胚胎第85天(E85)达到峰值,随后逐渐下降,并在出生后第3天(P3)后达到最低点。这与围产期异质性降低的发育“杯状”模型相符,并为其增添了更多细节。值得注意的是,皮层区域的差异表达基因数量仅为皮层下区域的三分之一,这一发现与之前证实的皮层内异质性降低的结果一致。通过将数据与已建立的人类和恒河猴发育数据集进行系统比对,实现了跨物种整合。比对结果支持了转录组水平上这种“上升-下降”轨迹的进化保守性,其最大异质性出现在围产期(猪胚胎第85天,对应人类原发性细胞死亡第194天),而出生后(猪出生后第3天附近,对应人类出生后第74天)的差异最小 (图 4h-i)。虽然人类和恒河猴数据集显示出类似的趋势,但猪脑图谱更高的围产期分辨率使得他们能够将区域异质性峰值精确定位到延长的E85时期(图 4h-i),从而为围产期神经发育提供了新的见解。除了转录动态之外,他们还系统地研究了发育不同时间点的区域间蛋白质组异质性。出乎意料的是,DEP的时间模式与转录组分化呈现出不同的轨迹:DEP的数量从胚胎第76天(E76)开始逐渐增加,并在胚胎第109天(E109)附近达到峰值,这一趋势与转录组和蛋白质组的测序深度差异无关。在食蟹猴蛋白质组数据集中也观察到了这种不同的动态变化,表明这种解耦的转录-翻译机制在进化上是保守的。功能分析进一步揭示,这些上调的DEP与GTP酶调节活性、囊泡组织和突触后致密区显著相关。
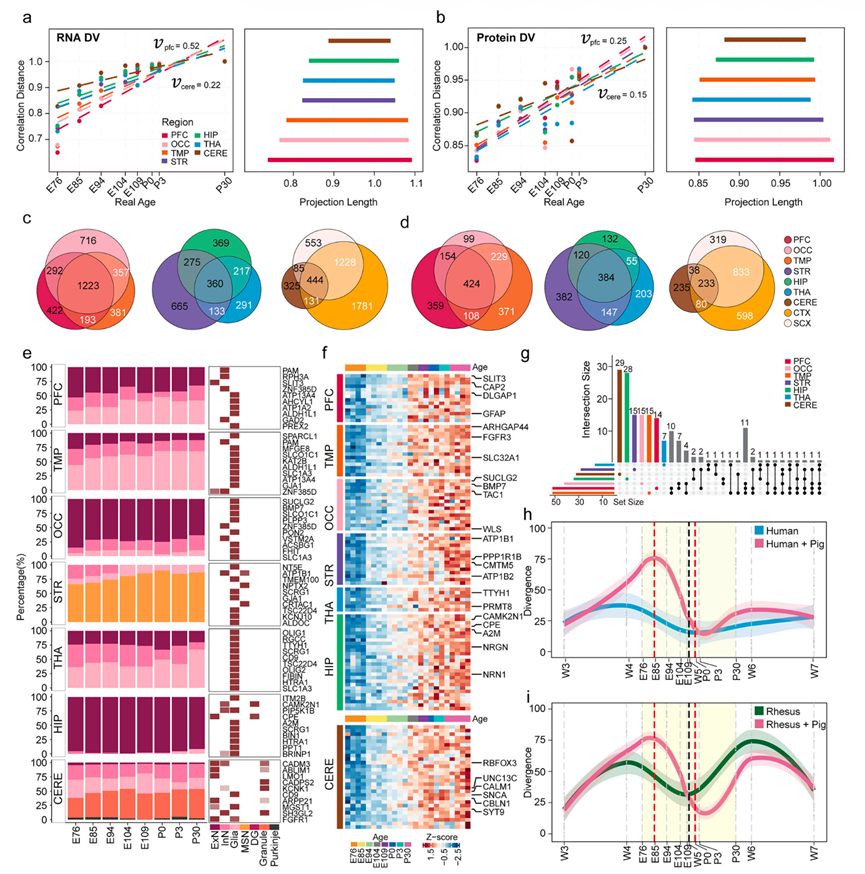
图4. 发育速度揭示了围产期的空间差异。
(a-b) 对不同脑区的相关距离(y 轴)与实际年龄(x 轴)进行线性回归分析,并按发育速度从下到上排列回归线的 y 轴投影长度。(c-d) 维恩图显示了不同脑区中发育速度基因群 (DVG)和发育速度预测点 (DVP)的重叠情况。(e) RNA-seq 数据的反卷积分析揭示了不同细胞类型在不同发育年龄和脑区中的比例分布(左),以及每个脑区前 10 个高置信度的细胞类型相关 DVG(颜色强度反映基因与细胞类型关联的强度)。(f) 热图展示了不同发育阶段区域特异性、高置信度细胞类型相关差异表达基因( DVG)的表达模式。(g) 所有高置信度细胞类型相关差异表达基因在各脑区的分布。(h-i) 通过差异表达分析和跨物种整合构建的全脑异质性时间曲线揭示了发育过程中动态的区域异质性模式。
05
整合的基因-蛋白质网络描绘了大脑发育的时空特征
虽然他们鉴定出了转录组-蛋白质组之间一致和不一致的关系,但仍尝试系统地分析跨脑区的7935个共检测到的基因-蛋白质对,以进一步研究它们的协调性。约25%的基因-蛋白质对表现出显著的表达相关性,其中70%在所有脑区均表现出保守的正向共变关系(图 5a-b)。功能富集分析表明,这些正相关的基因-蛋白质集中在轴突发育和突触组装通路中,而小脑特异性基因-蛋白质对的区域间共有率显著较低,突显了区域间调控优先级的差异。此外,他们还研究了转录因子(TF)与转录组以及RNA结合蛋白(RBP)与转录组之间的调控关系(图 5c-f)。为了最大限度地减少不必要的噪声影响,他们重点关注每个TF与其相关性最高的25%靶基因之间的调控关系。他们观察到相关系数呈双峰分布,表明TF可能具有双向调控作用,既包含激活也包含抑制(图 5c-d)。相比之下,RBP与其靶RNA之间的相关性在所有脑区均表现出显著的正调控倾向,其中60%的RBP-RNA对表现出正调控(图 5a-f),表明RBP是转录-翻译一致性的关键调节因子。
随后,加权基因共表达网络分析(WGCNA)将这些共检测到的基因-蛋白组合划分为11个组,包含69个模块(图 5g)。虽然许多模块(69个模块中的25个)表现出单一基因或蛋白的优势(主要成分的占比>95%),这与先前的转录-翻译不一致性观察结果一致,但大多数模块仍保持着基因和蛋白的可定量共现性(图 5g)。他们发现WGCNA分组和模块揭示了不同的时间或空间关联,例如G3组随着发育时间的推移表现出逐渐激活(正时间相关性)(图 5g),而G8组则表现出反向调控(负时间相关性)(图 5g)。区域特异性评估显示,ME26 和 ME62 是代表性模块,其核心分子主要在纹状体或海马中表达,包括PPP1R1B(纹状体)和ZBTB20(海马)。他们还对 WGCNA 组/模块的细胞成分进行了表征,发现 G8 和 G9 中神经元标记基因显著富集,尤其是在皮层神经元(ExN、InN)、海马齿状回神经元和纹状体 MSN 中(图 5g)。
此外,他们将WGCNA模块整合到一个四层调控轴中,构建了一个转录-翻译共调控网络。他们注意到许多模块与转录-翻译一致性、转录因子结合密度、蛋白质-蛋白质相互作用强度以及动态的区域间异质性模式均存在显著关联(图 5g)。尤其值得注意的是,载脂蛋白D(APOD)此前报道仅限于少突胶质细胞谱系细胞和脑内皮细胞,但在ME5中表现出基因和蛋白质的协同富集。时间序列分析显示APOD基因和蛋白质均呈进行性上调。为了进行更深入的表征,由于G3具有高度的转录-翻译一致性和显著的时空发育相关性(图 5g),因此优先选为模型调控枢纽。网络分析显示,ME5和ME7是G3内的关键节点,其中ME5富集于胶质细胞生成和脂质代谢过程,而ME7调控突触组装和投射神经元成熟。此外,枢纽基因分析证实,ME5包含经典的少突胶质细胞谱系标记物(MOG、MBP、MAG、PLP1),而ME7富集突触调节因子CNTNAP1和GABARAPL1。对ME5和ME7调控轴的进一步分析表明,共检测到的基因-蛋白表达谱在 E104 时同时激增,与先前确定的围产期过渡点相吻合(图3e-h)。在空间分布上,ME5 在皮层下区域和小脑中的表达显著高于大脑皮层,证实了非皮层区域神经胶质细胞成熟加速,并与已建立的神经胶质细胞生成时间线相符(图 5h)。此外,转录因子(TF)和RNA结合蛋白(RBP)关联分析表明,STAT3、TFEB和ITGB2等TF分别正向调控ME5和ME7中的基因转录,而PARS2、SEPTN4、SARS2和SUPV3L1等RBP则通过转录后调控促进蛋白质翻译(图 5h)。
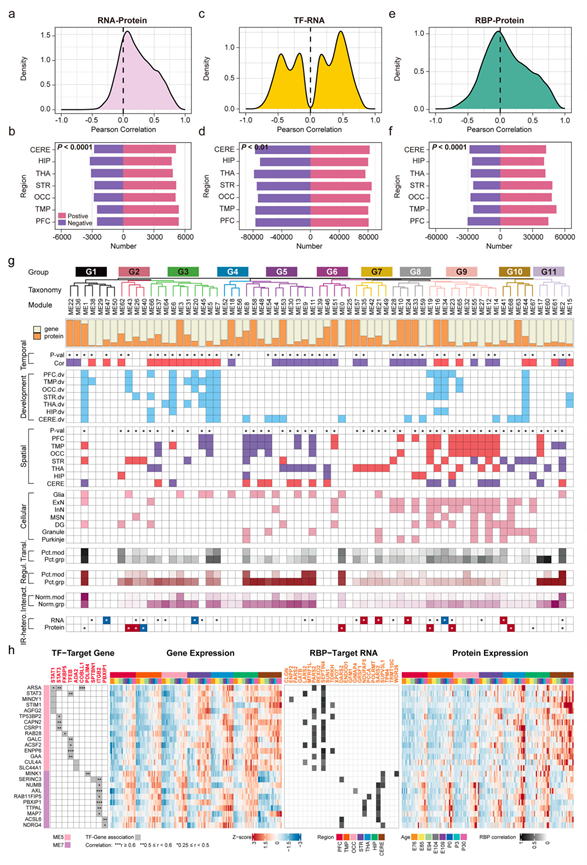
图5. 整合基因-蛋白质共表达网络分析。
(a) 分别使用RNA-seq数据和蛋白质组数据计算共检测到的基因-蛋白质对之间相关系数的密度分布。(b) 定量脑区中共检测到的基因(RNA-seq数据)-蛋白质(蛋白质组数据)对的相关性。(c) 分别使用蛋白质组数据和RNA-seq数据计算的转录因子(TF)与其前25%靶基因之间相关系数的密度分布。(d) 定量脑区中转录因子(蛋白质组数据)与靶基因的相关性。(e) 用蛋白质组数据计算的RNA结合蛋白(RBP)与其靶RNA之间相关系数的密度分布。(f) 定量脑区中RBP(蛋白质组数据)与靶RNA的相关性。(g) WGCNA基于特征基因以及各模块内基因和蛋白质的组成分布,鉴定出69个模块,并将其聚类为11个组。(h)相关性分析。
06
围产期发育过程中脑疾病风险基因的动态分析
围产期发育紊乱会增加日后患病的易感性,但其分子机制仍未得到充分阐明。通过整合猪围产期和人类发育转录组的跨物种数据,他们克服了人类神经发育研究中的一个主要障碍,从而能够对疾病相关的分子改变进行最全面的生命周期探索(图 6a)。在脑发育的第一阶段(早期胎儿期),他们对相关疾病的表达模式进行了聚类分析,其中精神疾病聚类在一起,神经胶质瘤和与皮质畸形相关的疾病聚类在一起。随着发育的进行,疾病风险基因相关性的组织逐渐破坏,这种破坏程度在E76-E94期间达到峰值(对应于人类原发性纤毛运动障碍183-206期);这种紊乱随后在胚胎发育第104天至出生后第3天(E104-P3,相当于人类原发性细胞死亡257天至出生后第74天)期间逆转,转变为一种持续到成年的“泛相关”模式。反过来,他们利用成年期疾病风险基因的表达构建了聚类框架,然后将所有发育阶段投影到该框架上。他们观察到了类似的模式,即胚胎发育第76天至第94天(E76-E94)阶段和胚胎发育第104天至出生后第3天(E104-P3)阶段之间存在明显的基因分离,其中E104天至出生后第3天(E104-P3)的表达谱与成年期的表达谱非常相似。此外,疾病相关风险基因集的终生表达动态也始终支持这一模式。综合来看,这些分析方法一致表明,猪脑数据无缝衔接了人类早期胎儿期到成年期疾病基因表达模式的演变动态,并且猪脑在 E94–E104 期间经历了一次重大的发育转变,E104-P3 阶段已经表现出与成年期(人类)高度相似的组织结构。
随后,他们对上述11个WGCNA分组中的疾病风险基因进行了富集分析,以进一步了解围产期神经发育与疾病发病机制之间的关联。他们发现大多数分组(G1、G5、G6、G8和G9)与特定疾病集存在显著相关性,其中G5组显示出更广泛的疾病特异性关联(图 6b)。此外,他们发现ASD(例如SFARI.Score1)、无脑回畸形和多小脑回畸形也显示出更广泛的分组富集关联(图 6b)。接下来,重点关注了四个关联性最显著的疾病-分组组合:无脑回畸形与G1组、发育迟缓(DD)和ASD(SFARI.Score1)与G5组以及ASD(SFARI.Score1)与G9组。他们构建了这些风险基因在其各自组内的分子互作网络(图 6c)。值得注意的是,在所有分析中,疾病风险基因均作为网络枢纽发挥作用,其中特征明确的ASD风险基因(例如SYNGAP1、GABRB3)在ASD网络分析(SFARI.Score1)中占据中心节点位置,并与G9进行了比较(图 6c)。他们构建了目标基因组的加权邻接矩阵,并量化了各个节点的连接强度。统计分析表明,这些组内富集的神经发育疾病核心风险基因和所有神经发育疾病风险基因的网络连接强度显著高于组平均水平,提示这些风险基因可能在共表达网络中发挥关键的枢纽作用(图 6d-g)。总而言之,这些发现强调了围产期脑发育与疾病进展之间的密切联系,为理解神经发育障碍的分子变化提供了新的方向。
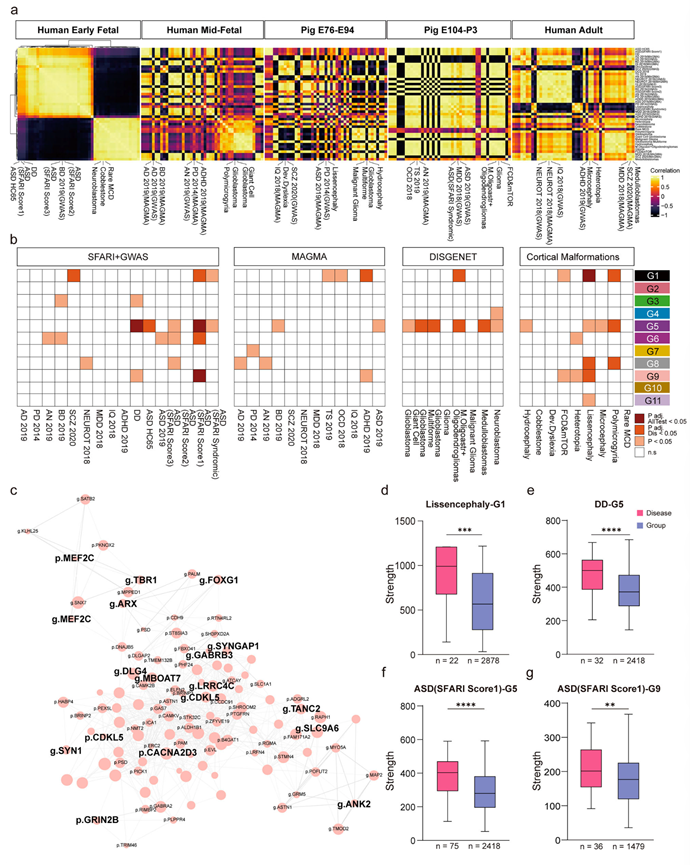
图6. 转录组/蛋白质组谱中脑疾病风险基因的动态表征。
(a) 热图展示了整合的人-猪转录组在五个发育阶段(人类早期胎儿、人类中期胎儿、猪 E76-E94、猪 E104-P3 和人类成年期)中疾病风险基因集的特征基因的 Pearson 相关性。(b) 疾病风险基因集在 WGCNA 分组中的富集情况。(c) 由ASD(SFARI Score1)风险基因与G9中所有分子的交集构建的分子相互作用网络(方法)。(d-g) 对每个WGCNA定义的组内疾病相关风险分子和非风险分子之间的连接强度进行统计比较。
+ + + + + + + + + + +
结 论
本研究分析了336个巴马小型猪围产期样本在多个时间点和脑区的转录组和蛋白质组数据,揭示了其时空组织结构以及基因和蛋白质之间的一致性。他们发现猪是理想的大型动物模型,其与人类的相似性更高,与猴子相似,且优于小鼠和其他物种。同时,本研究还发现猪的围产期是连接人类早期产前到产后分子轨迹的过渡枢纽,揭示了围产期存在一个泛区域的发育拐点(E94-E104),表现为与突触发生和胶质细胞生成相关的基因和蛋白质表达水平的变化。在进一步分析猪脑各区域的发育速度时,本研究发现区域异质性在围产期呈现出显著的同步上升-下降模式。此外,脑部疾病风险基因在猪脑中也表现出从围产期向成体样组织结构的转变,这使得该多组学图谱成为解码神经发育障碍发病机制的转化框架。
+ + + + +



